

ATP13A3 variants promote pulmonary arterial hypertension by disrupting polyamine transport
- PMID: 38626311
- PMCID: PMC11135649
- DOI: 10.1093/cvr/cvae068
ATP13A3 variants promote pulmonary arterial hypertension by disrupting polyamine transport
Abstract
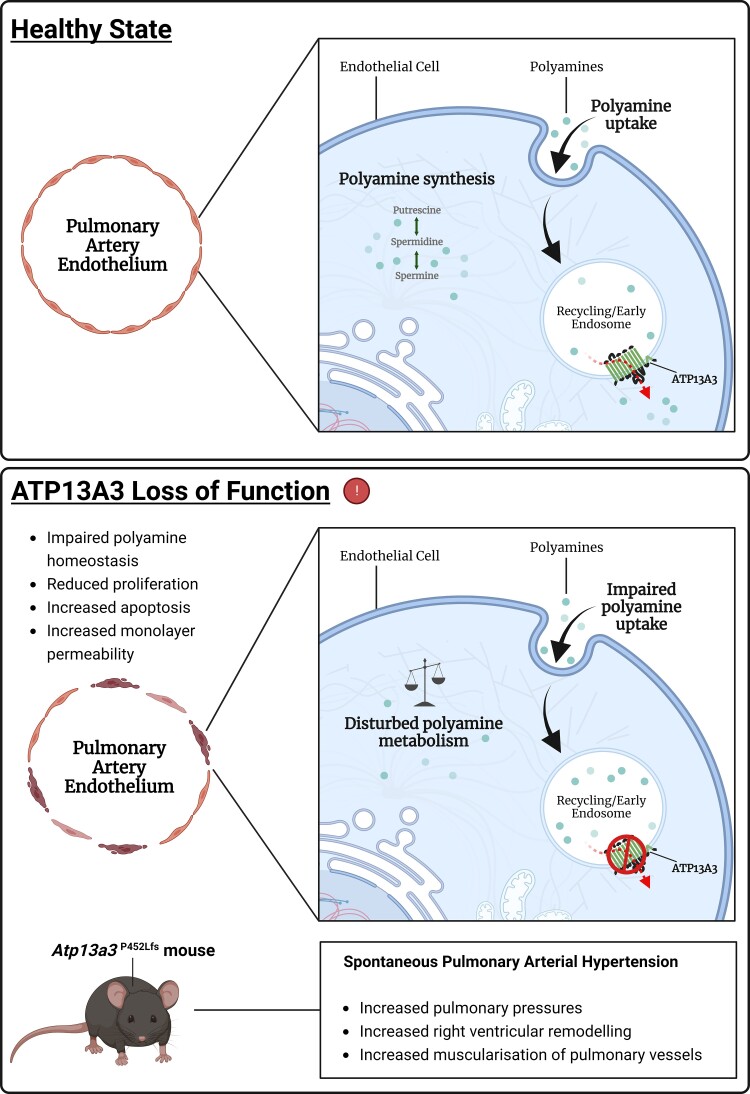
Aims: Potential loss-of-function variants of ATP13A3, the gene encoding a P5B-type transport ATPase of undefined function, were recently identified in patients with pulmonary arterial hypertension (PAH). ATP13A3 is implicated in polyamine transport but its function has not been fully elucidated. In this study, we sought to determine the biological function of ATP13A3 in vascular endothelial cells (ECs) and how PAH-associated variants may contribute to disease pathogenesis.
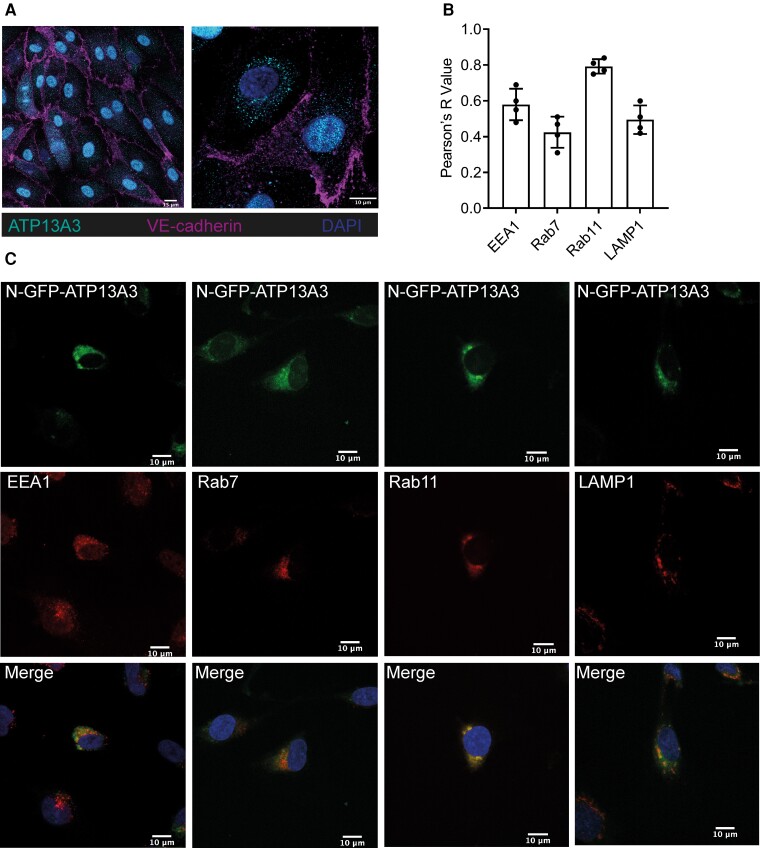
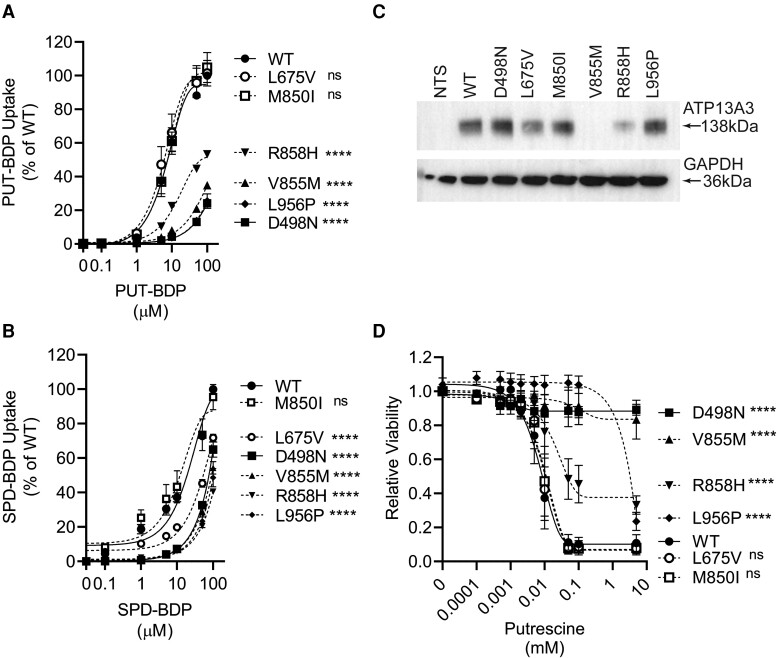
Methods and results: We studied the impact of ATP13A3 deficiency and overexpression in EC models [human pulmonary ECs, blood outgrowth ECs (BOECs), and human microvascular EC 1], including a PAH patient-derived BOEC line harbouring an ATP13A3 variant (LK726X). We also generated mice harbouring an Atp13a3 variant analogous to a human disease-associated variant to establish whether these mice develop PAH. ATP13A3 localized to the recycling endosomes of human ECs. Knockdown of ATP13A3 in ECs generally reduced the basal polyamine content and altered the expression of enzymes involved in polyamine metabolism. Conversely, overexpression of wild-type ATP13A3 increased polyamine uptake. Functionally, loss of ATP13A3 was associated with reduced EC proliferation, increased apoptosis in serum starvation, and increased monolayer permeability to thrombin. The assessment of five PAH-associated missense ATP13A3 variants (L675V, M850I, V855M, R858H, and L956P) confirmed loss-of-function phenotypes represented by impaired polyamine transport and dysregulated EC function. Furthermore, mice carrying a heterozygous germline Atp13a3 frameshift variant representing a human variant spontaneously developed a PAH phenotype, with increased pulmonary pressures, right ventricular remodelling, and muscularization of pulmonary vessels.
Conclusion: We identify ATP13A3 as a polyamine transporter controlling polyamine homeostasis in ECs, a deficiency of which leads to EC dysfunction and predisposes to PAH. This suggests a need for targeted therapies to alleviate the imbalances in polyamine homeostasis and EC dysfunction in PAH.
Keywords: ATP13A3; Polyamines; Pulmonary arterial hypertension.
© The Author(s) 2024. Published by Oxford University Press on behalf of the European Society of Cardiology.
Conflict of interest statement
Conflict of interest: P.D.U. and N.W.M. have published US (US10336800) and EU (EP3166628B1) patents entitled: ‘Therapeutic Use of Bone Morphogenetic Proteins’. All other authors declare no competing interests.
Figures





References
-
- Long L, Ormiston ML, Yang X, Southwood M, Gräf S, Machado RD, Mueller M, Kinzel B, Yung LM, Wilkinson JM, Moore SD, Drake KM, Aldred MA, Yu PB, Upton PD, Morrell NW. Selective enhancement of endothelial BMPR-II with BMP9 reverses pulmonary arterial hypertension. Nat Med 2015;21:777–785. - PMC - PubMed
-
- Lane KB, Machado RD, Pauciulo MW, Thomson JR, Phillips JA III, Loyd JE, Nichols WC, Trembath RC. Heterozygous germline mutations in BMPR2, encoding a TGF-beta receptor, cause familial primary pulmonary hypertension. The International PPH Consortium. Nat Genet 2000;26:81–84. - PubMed
-
- Deng Z, Morse JH, Slager SL, Cuervo N, Moore KJ, Venetos G, Kalachikov S, Cayanis E, Fischer SG, Barst RJ, Hodge SE, Knowles JA. Familial primary pulmonary hypertension (gene PPH1) is caused by mutations in the bone morphogenetic protein receptor-II gene. Am J Hum Genet 2000;67:737–744. - PMC - PubMed
-
- Southgate L, Machado RD, Gräf S, Morrell NW. Molecular genetic framework underlying pulmonary arterial hypertension. Nat Rev Cardiol 2020;17:85–95. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
