

The physiological interactome of TCR-like antibody therapeutics in human tissues
- PMID: 38627373
- PMCID: PMC11021511
- DOI: 10.1038/s41467-024-47062-5
The physiological interactome of TCR-like antibody therapeutics in human tissues
Abstract
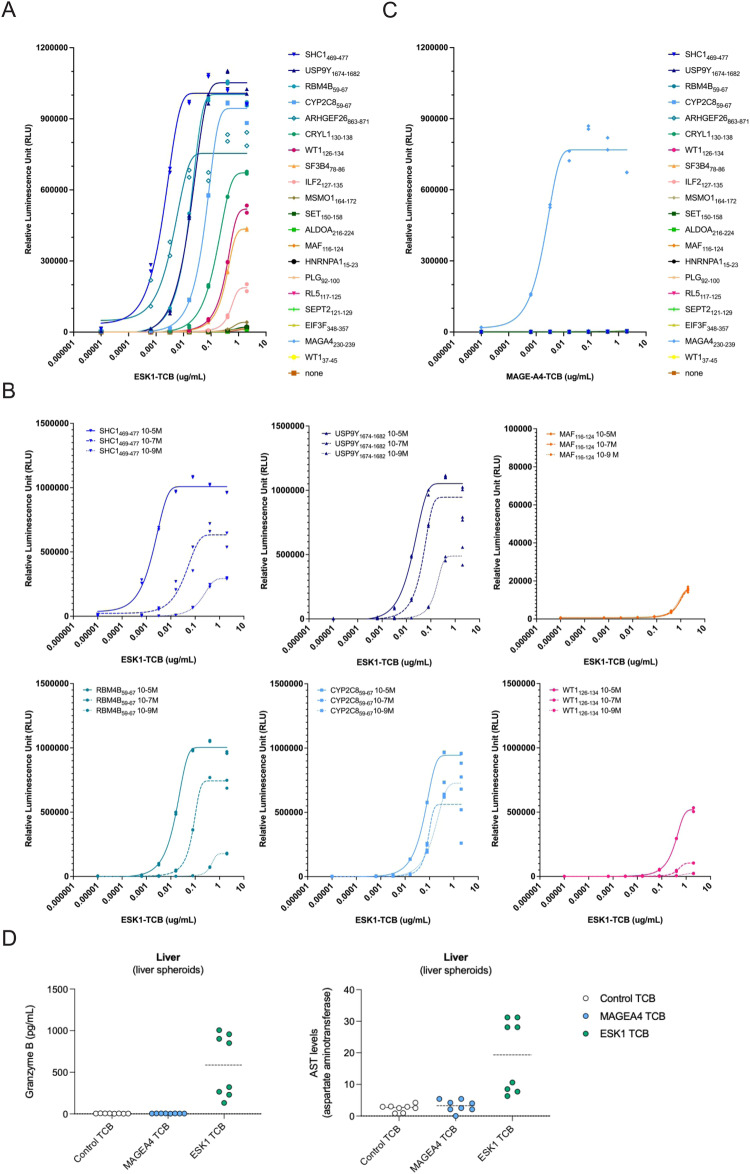
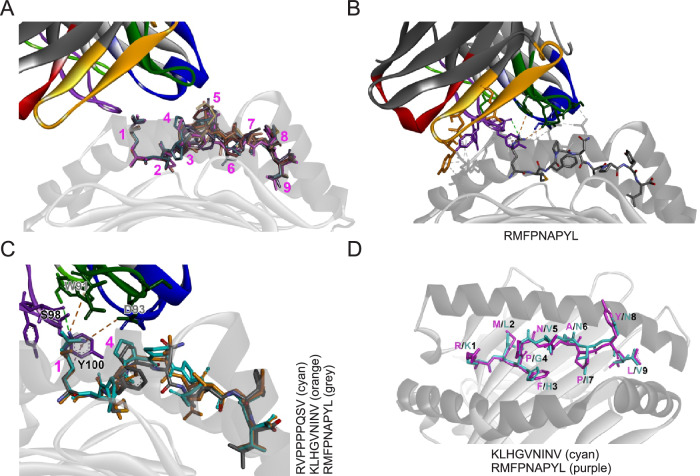
Selective binding of TCR-like antibodies that target a single tumour-specific peptide antigen presented by human leukocyte antigens (HLA) is the absolute prerequisite for their therapeutic suitability and patient safety. To date, selectivity assessment has been limited to peptide library screening and predictive modeling. We developed an experimental platform to de novo identify interactomes of TCR-like antibodies directly in human tissues using mass spectrometry. As proof of concept, we confirm the target epitope of a MAGE-A4-specific TCR-like antibody. We further determine cross-reactive peptide sequences for ESK1, a TCR-like antibody with known off-target activity, in human liver tissue. We confirm off-target-induced T cell activation and ESK1-mediated liver spheroid killing. Off-target sequences feature an amino acid motif that allows a structural groove-coordination mimicking that of the target peptide, therefore allowing the interaction with the engager molecule. We conclude that our strategy offers an accurate, scalable route for evaluating the non-clinical safety profile of TCR-like antibody therapeutics prior to first-in-human clinical application.
© 2024. The Author(s).
Conflict of interest statement
All authors listed with Roche Pharma affiliation are directly employed by Roche Pharma. T.W., P.U., and C.K. are inventors on related patents. P.U. and C.K. declare stock ownership. N.T. has been a paid consultant to Roche Pharma. The remaining authors declare no competing interests.
Figures



References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
