

Complement C5a Receptor Signaling Alters Stress Responsiveness and Modulates Microglia Following Chronic Stress Exposure
- PMID: 38628385
- PMCID: PMC11019103
- DOI: 10.1016/j.bpsgos.2024.100306
Complement C5a Receptor Signaling Alters Stress Responsiveness and Modulates Microglia Following Chronic Stress Exposure
Abstract
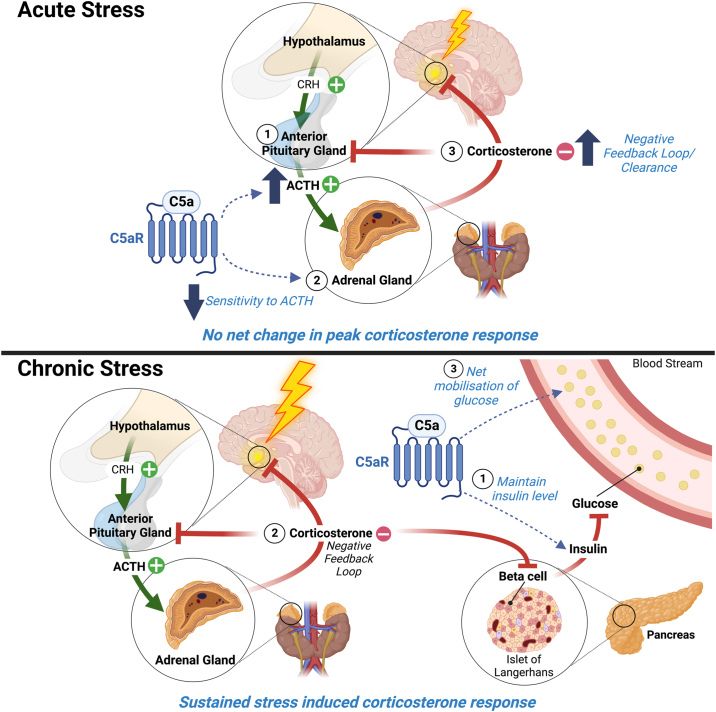
Background: Accumulating evidence underscores the pivotal role of heightened inflammation in the pathophysiology of stress-related diseases, but the underlying mechanisms remain elusive. The complement system, a key effector of the innate immune system, produces the C5-cleaved activation product C5a upon activation, initiating inflammatory responses through the canonical C5a receptor 1 (C5aR1). While C5aR1 is expressed in stress-responsive brain regions, its role in stress responsiveness remains unknown.
Methods: To investigate C5a-C5aR1 signaling in stress responses, mice underwent acute and chronic stress paradigms. Circulating C5a levels and messenger RNA expression of C5aR1 in the hippocampus and adrenal gland were measured. C5aR1-deficient mice were used to elucidate the effects of disrupted C5a-C5aR1 signaling across behavioral, hormonal, metabolic, and inflammation parameters.
Results: Chronic restraint stress elevated circulating C5a levels while reducing C5aR1 messenger RNA expression in the hippocampus and adrenal gland. Notably, the absence of C5aR1 signaling enhanced adrenal sensitivity to adrenocorticotropic hormone, concurrently reducing pituitary adrenocorticotropic hormone production and enhancing the response to acute stress. C5aR1-deficient mice exhibited attenuated reductions in locomotor activity and body weight under chronic stress. Additionally, these mice displayed increased glucocorticoid receptor sensitivity and disrupted glucose and insulin homeostasis. Chronic stress induced an increase in C5aR1-expressing microglia in the hippocampus, a response mitigated in C5aR1-deficient mice.
Conclusions: C5a-C5aR1 signaling emerges as a key metabolic regulator during stress, suggesting that complement activation and dysfunctional C5aR1 signaling may contribute to neuroinflammatory phenotypes in stress-related disorders. The results advocate for further exploration of complement C5aR1 as a potential therapeutic target for stress-related conditions.
Keywords: C5a receptor; Complement system; Corticosterone; Glia; Hippocampus; Neuroinflammation.
Plain language summary
How the immune system, particularly the complement system, influences responses to stress has not been fully clear. In this study, we focus on C5a-C5aR1 signaling, a part of the immune system, and found that it significantly affects stress-related reactions in mice. In chronic stress, we observed increased inflammation, altered hormonal responses, and disrupted metabolic regulation. Mice lacking C5aR1 showed reduced stress-induced behavioral changes, indicating that this receptor may play a vital role in modulating the stress response. Understanding these immune mechanisms sheds light on stress-related disorders and may open avenues for therapeutic interventions.
© 2024 The Authors.
Figures




References
-
- Horowitz M.A., Zunszain P.A. Neuroimmune and neuroendocrine abnormalities in depression: Two sides of the same coin. Ann N Y Acad Sci. 2015;1351:68–79. - PubMed
-
- Pape K., Tamouza R., Leboyer M., Zipp F. Immunoneuropsychiatry - Novel perspectives on brain disorders. Nat Rev Neurol. 2019;15:317–328. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
