

Modulation of peroxisomal import by the PEX13 SH3 domain and a proximal FxxxF binding motif
- PMID: 38632234
- PMCID: PMC11024197
- DOI: 10.1038/s41467-024-47605-w
Modulation of peroxisomal import by the PEX13 SH3 domain and a proximal FxxxF binding motif
Abstract
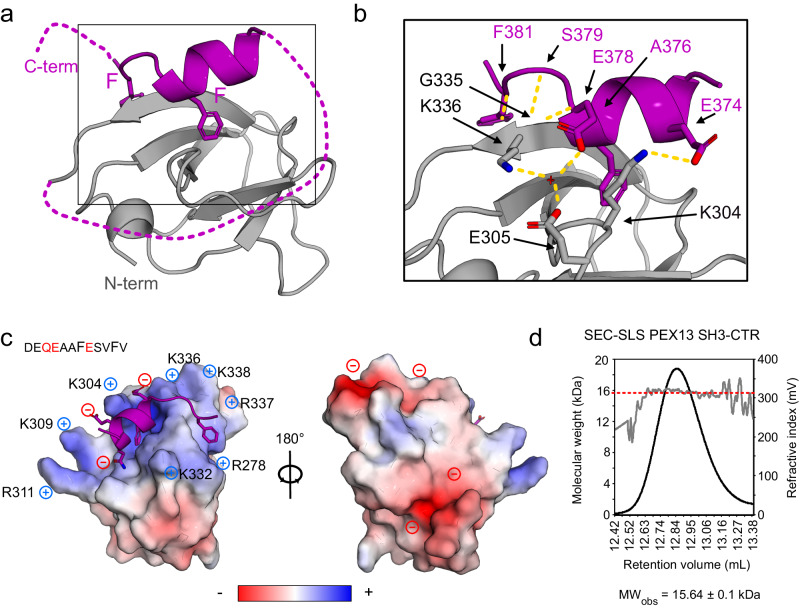
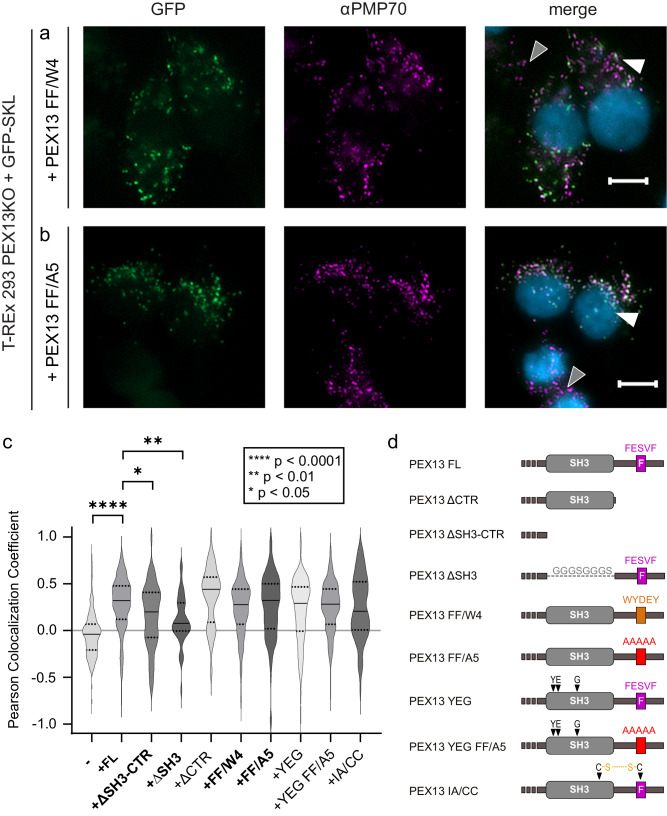
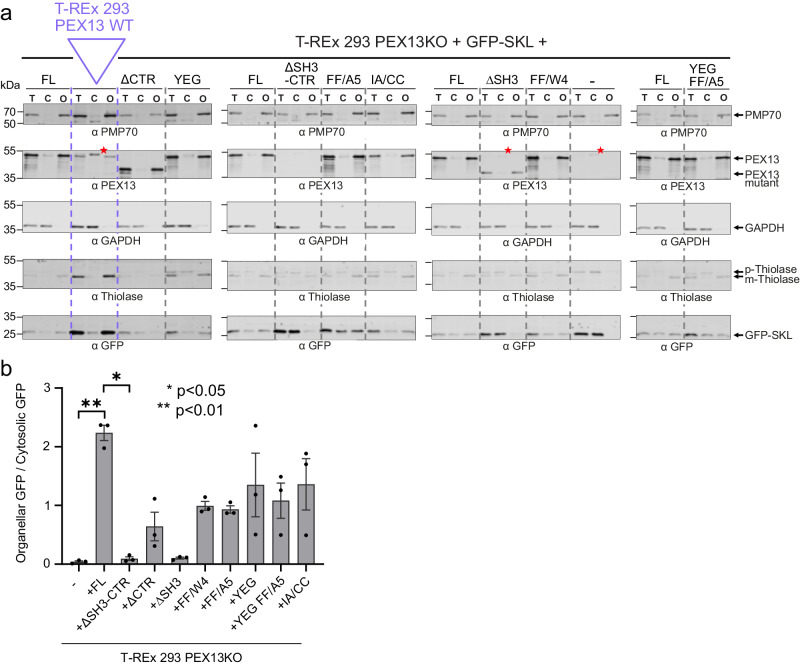
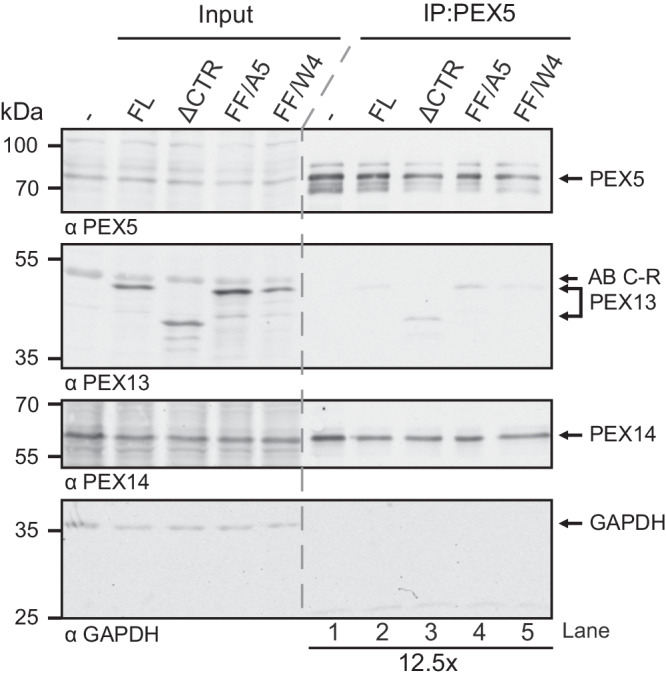
Import of proteins into peroxisomes depends on PEX5, PEX13 and PEX14. By combining biochemical methods and structural biology, we show that the C-terminal SH3 domain of PEX13 mediates intramolecular interactions with a proximal FxxxF motif. The SH3 domain also binds WxxxF peptide motifs in the import receptor PEX5, demonstrating evolutionary conservation of such interactions from yeast to human. Strikingly, intramolecular interaction of the PEX13 FxxxF motif regulates binding of PEX5 WxxxF/Y motifs to the PEX13 SH3 domain. Crystal structures reveal how FxxxF and WxxxF/Y motifs are recognized by a non-canonical surface on the SH3 domain. The PEX13 FxxxF motif also mediates binding to PEX14. Surprisingly, the potential PxxP binding surface of the SH3 domain does not recognize PEX14 PxxP motifs, distinct from its yeast ortholog. Our data show that the dynamic network of PEX13 interactions with PEX5 and PEX14, mediated by diaromatic peptide motifs, modulates peroxisomal matrix import.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
-
- Fujiki Y, Lazarow PB. Post-translational import of fatty acyl-CoA oxidase and catalase into peroxisomes of rat liver in vitro. J. Biol. Chem. 1985;260:5603–5609. - PubMed
-
- Erdmann R, Veenhuis M, Kunau WH. Peroxisomes: Organelles at the crossroads. Trends Cell Biol. 1997;7:400–407. - PubMed
-
- Wanders RJ, Waterham HR. Biochemistry of mammalian peroxisomes revisited. Ann. Rev. Biochem. 2006;75:295–332. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
