

Control of working memory by phase-amplitude coupling of human hippocampal neurons
- PMID: 38632400
- PMCID: PMC11078732
- DOI: 10.1038/s41586-024-07309-z
Control of working memory by phase-amplitude coupling of human hippocampal neurons
Abstract
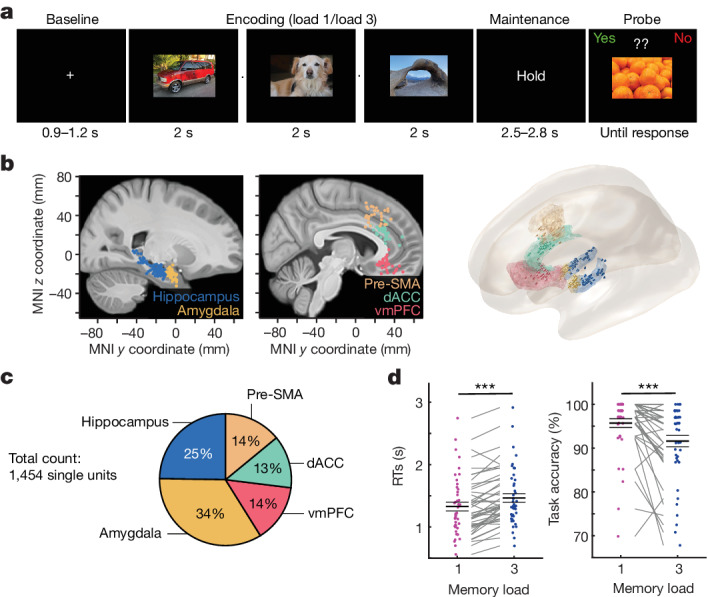
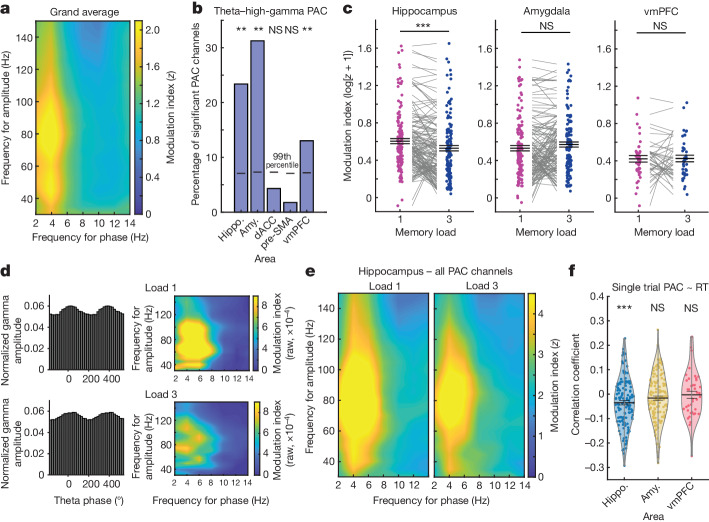
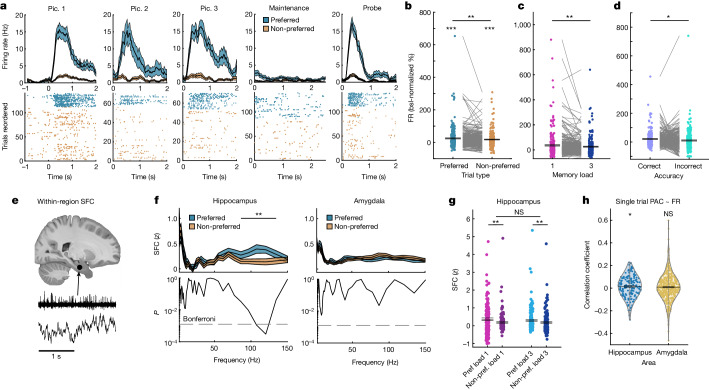
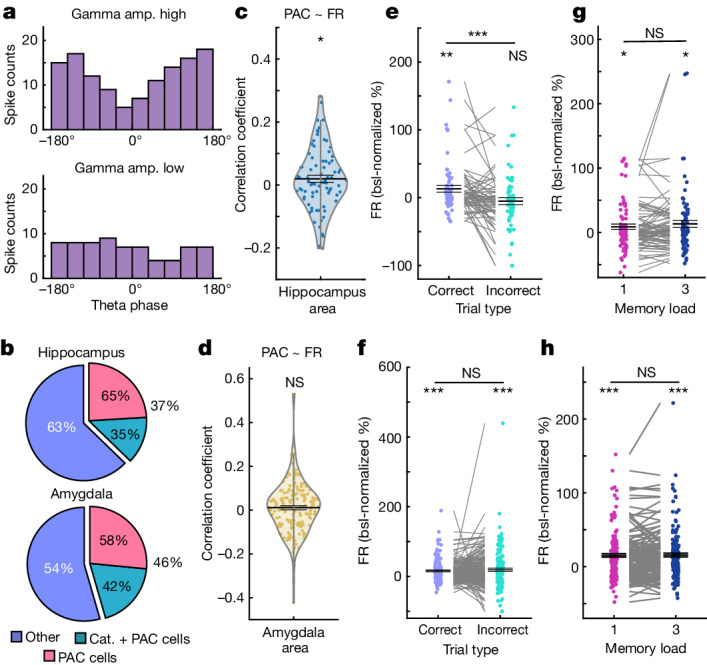
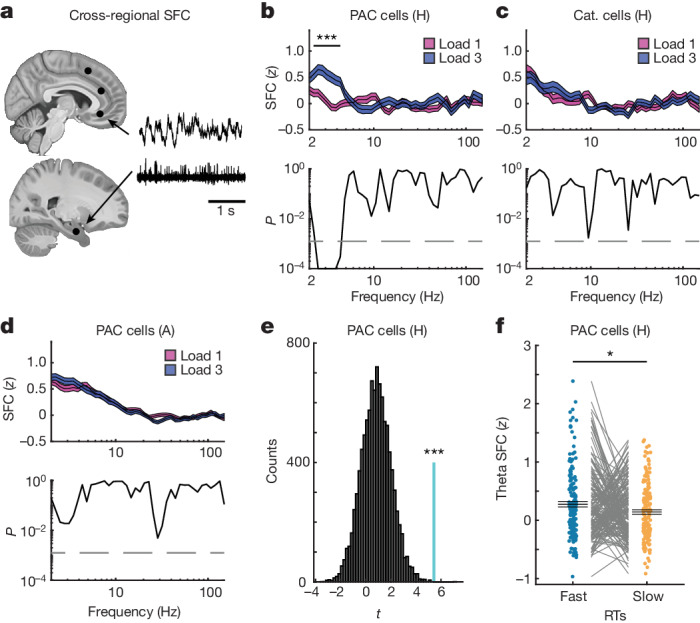
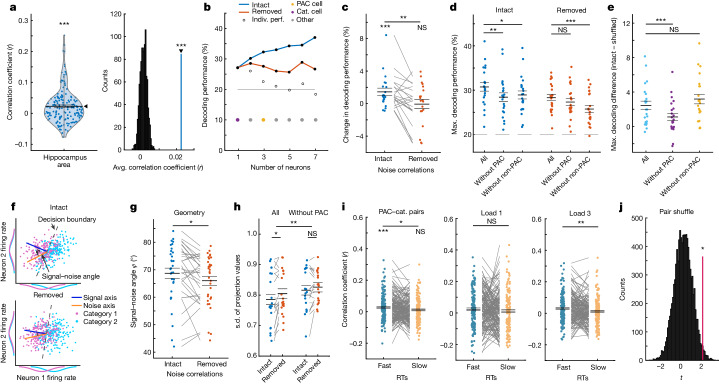
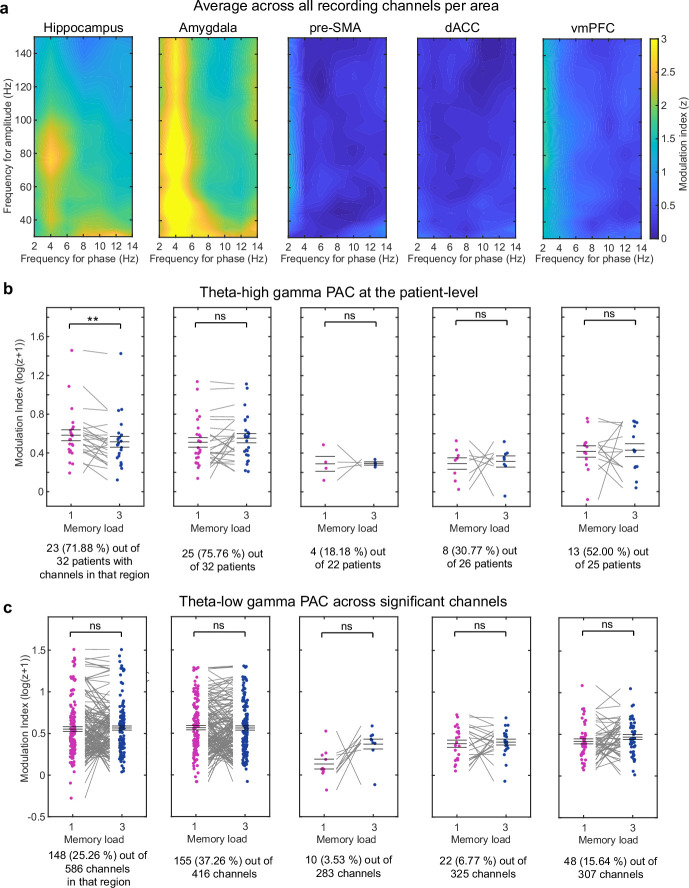
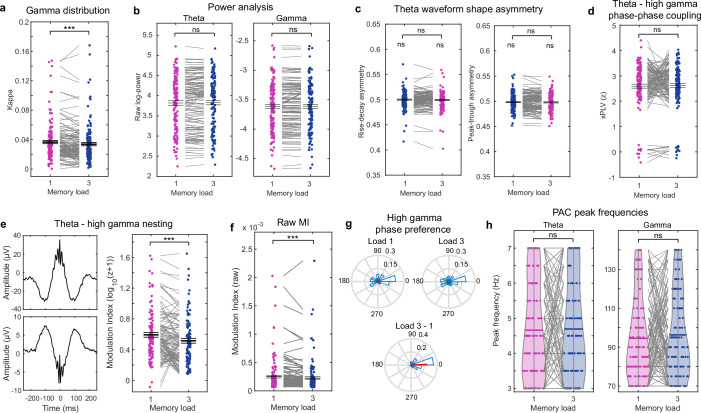
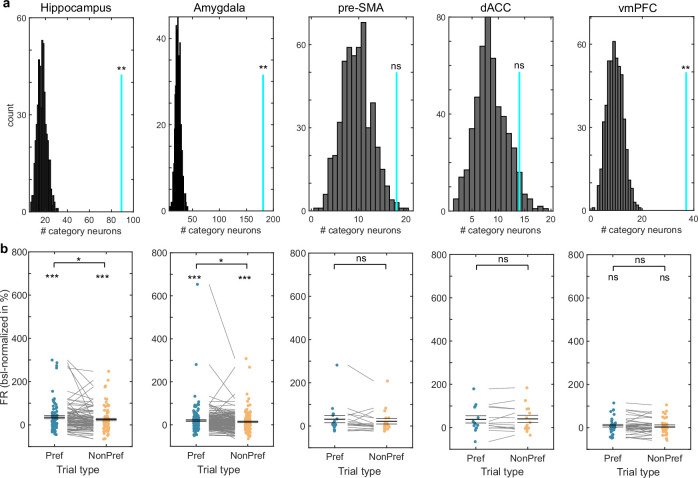
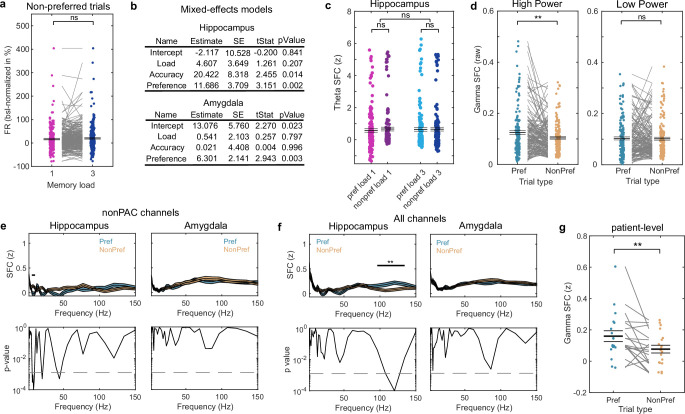
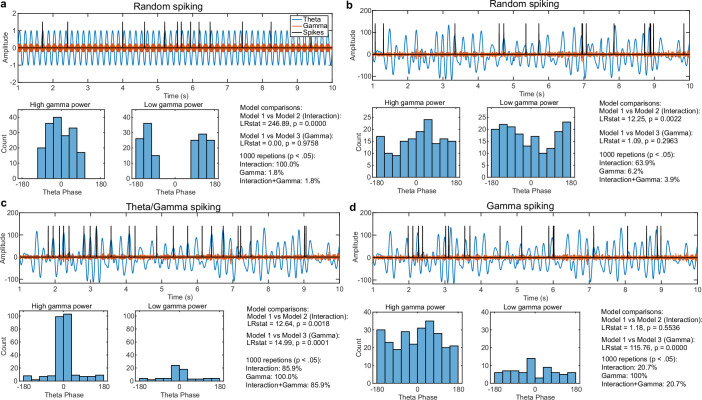
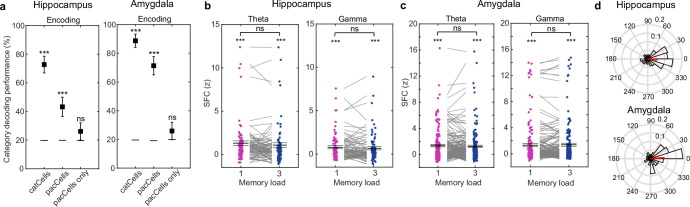
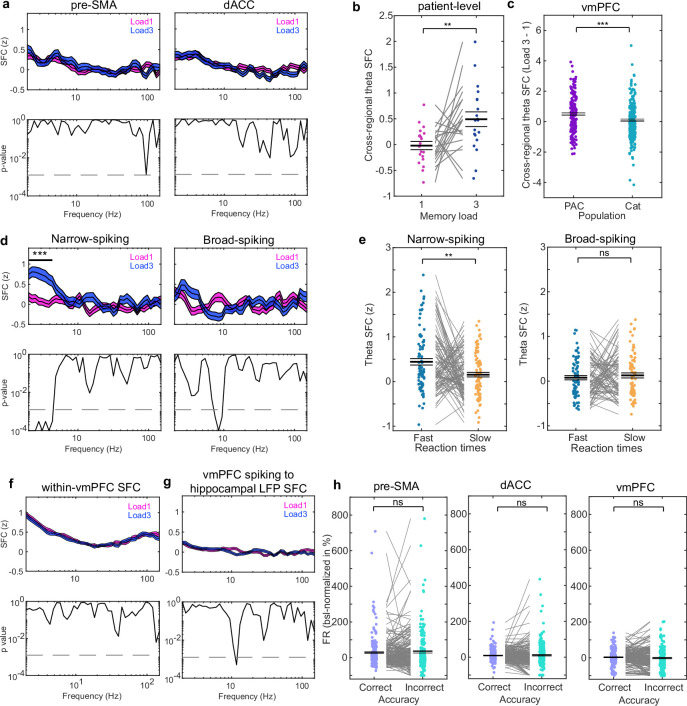
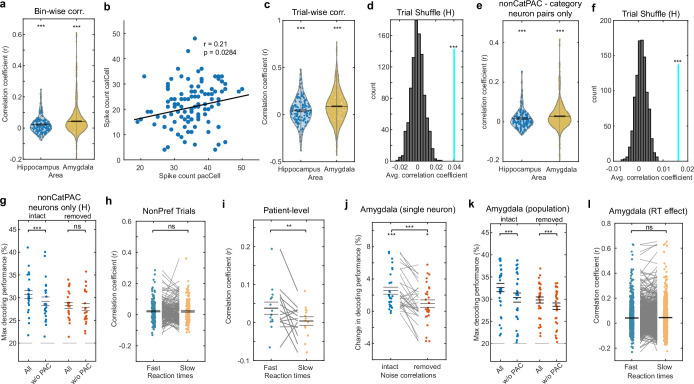
Retaining information in working memory is a demanding process that relies on cognitive control to protect memoranda-specific persistent activity from interference1,2. However, how cognitive control regulates working memory storage is unclear. Here we show that interactions of frontal control and hippocampal persistent activity are coordinated by theta-gamma phase-amplitude coupling (TG-PAC). We recorded single neurons in the human medial temporal and frontal lobe while patients maintained multiple items in their working memory. In the hippocampus, TG-PAC was indicative of working memory load and quality. We identified cells that selectively spiked during nonlinear interactions of theta phase and gamma amplitude. The spike timing of these PAC neurons was coordinated with frontal theta activity when cognitive control demand was high. By introducing noise correlations with persistently active neurons in the hippocampus, PAC neurons shaped the geometry of the population code. This led to higher-fidelity representations of working memory content that were associated with improved behaviour. Our results support a multicomponent architecture of working memory1,2, with frontal control managing maintenance of working memory content in storage-related areas3-5. Within this framework, hippocampal TG-PAC integrates cognitive control and working memory storage across brain areas, thereby suggesting a potential mechanism for top-down control over sensory-driven processes.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Control of working memory maintenance by theta-gamma phase amplitude coupling of human hippocampal neurons.bioRxiv [Preprint]. 2023 Apr 7:2023.04.05.535772. doi: 10.1101/2023.04.05.535772. bioRxiv. 2023. Update in: Nature. 2024 May;629(8011):393-401. doi: 10.1038/s41586-024-07309-z. PMID: 37066145 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
