

Tracing genetic diversity captures the molecular basis of misfolding disease
- PMID: 38637533
- PMCID: PMC11026414
- DOI: 10.1038/s41467-024-47520-0
Tracing genetic diversity captures the molecular basis of misfolding disease
Abstract
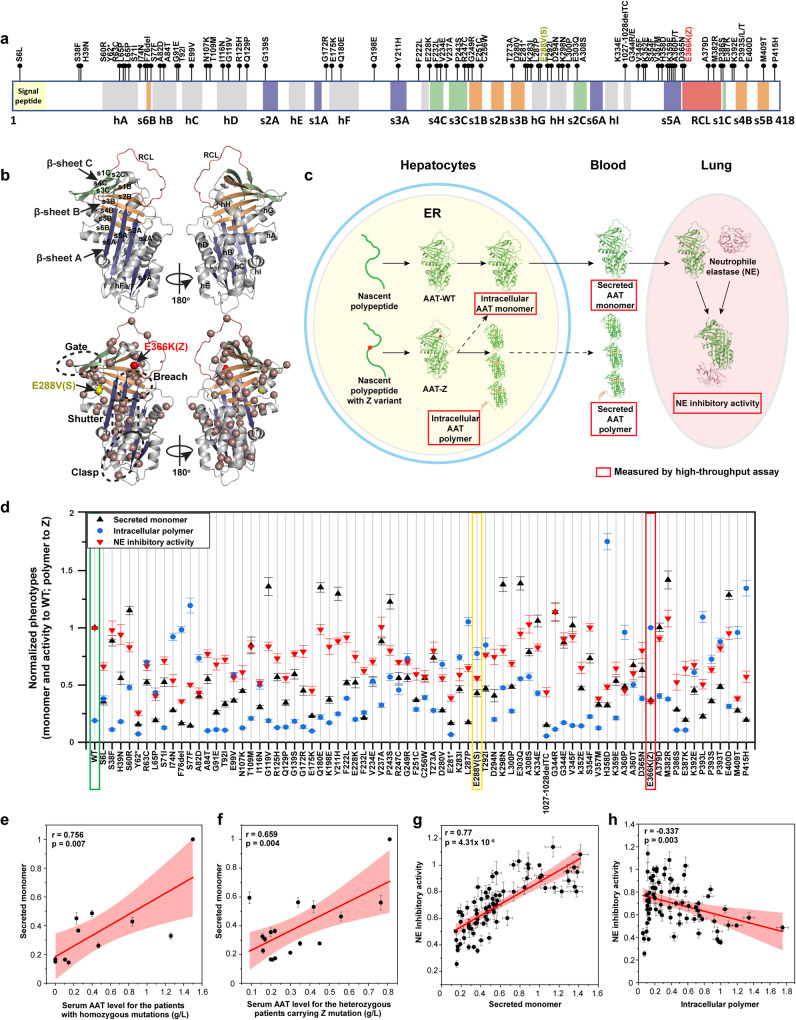
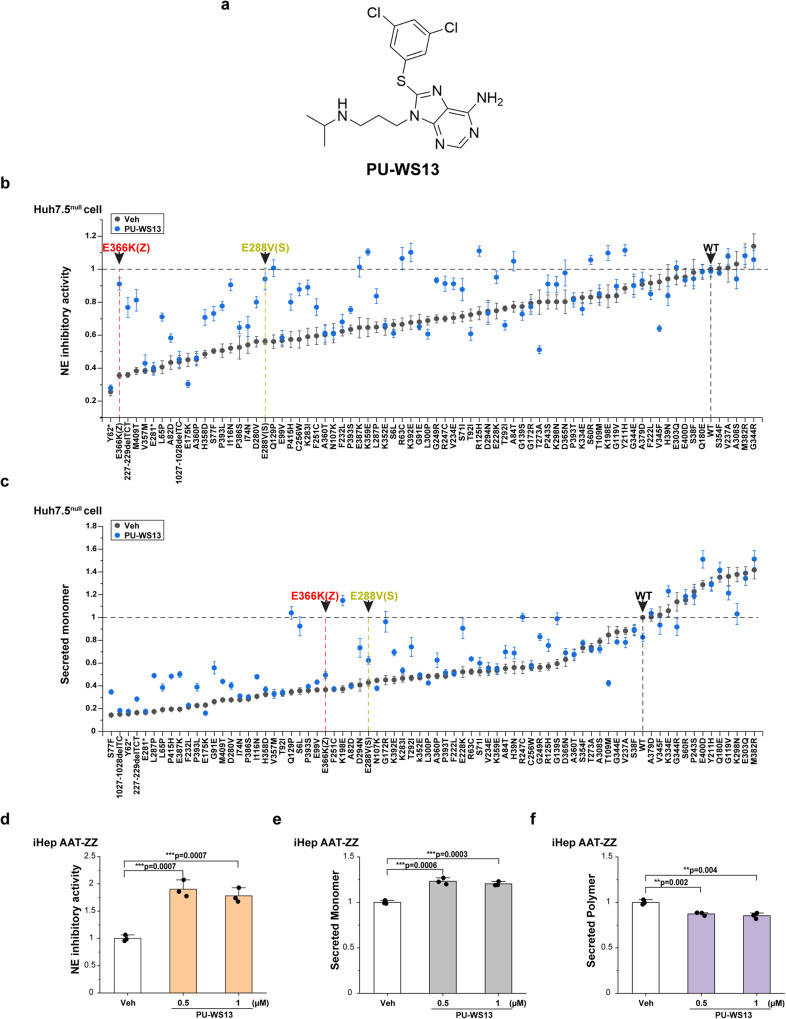
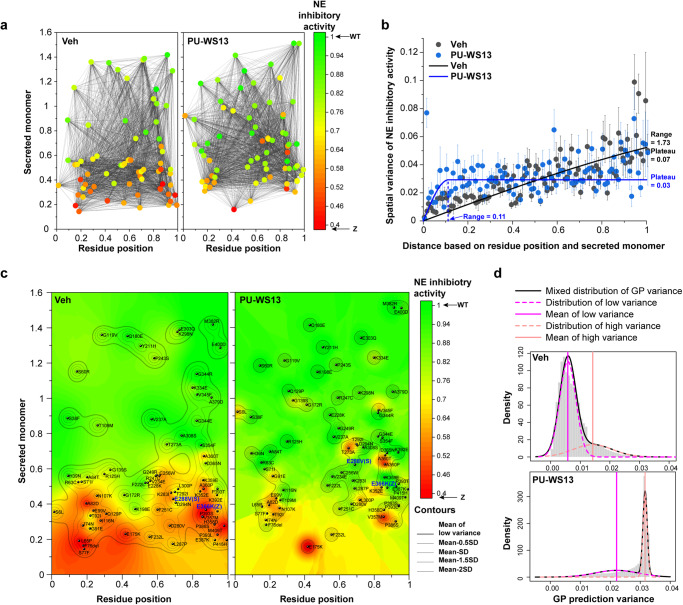
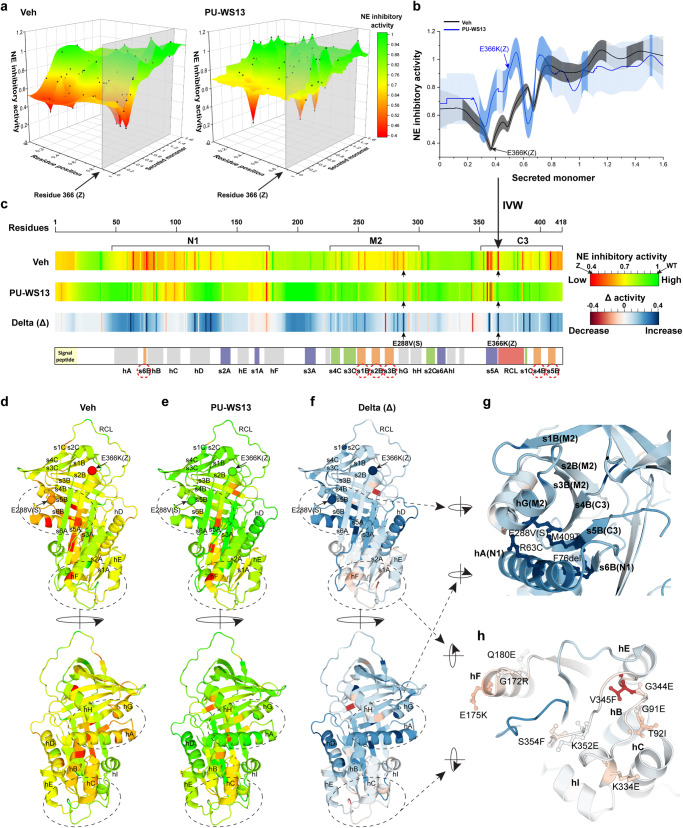
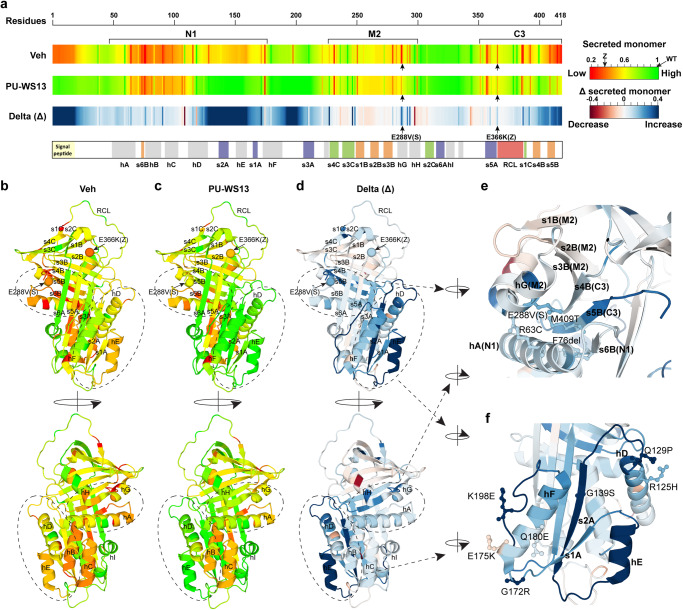
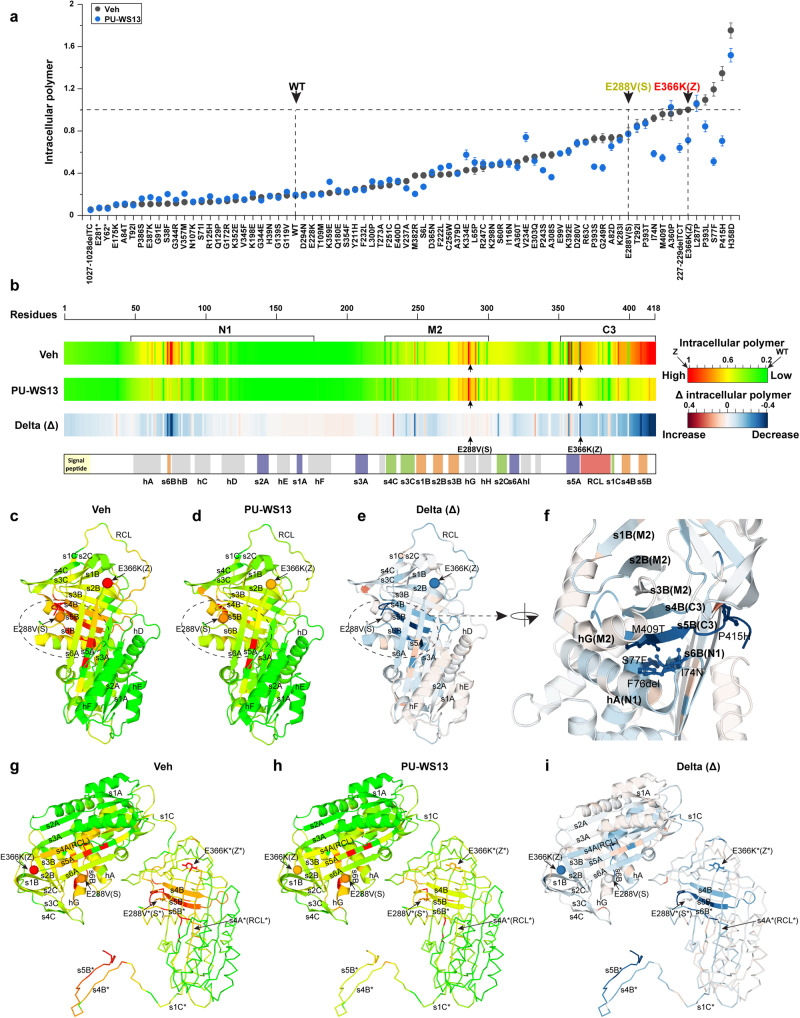
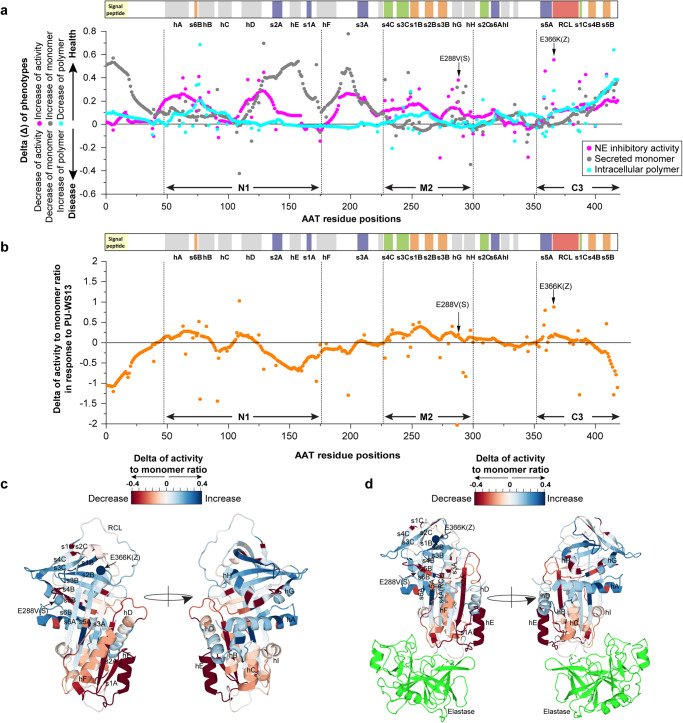
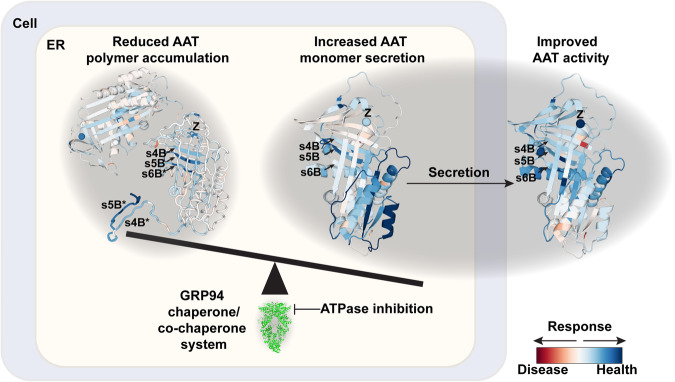
Genetic variation in human populations can result in the misfolding and aggregation of proteins, giving rise to systemic and neurodegenerative diseases that require management by proteostasis. Here, we define the role of GRP94, the endoplasmic reticulum Hsp90 chaperone paralog, in managing alpha-1-antitrypsin deficiency on a residue-by-residue basis using Gaussian process regression-based machine learning to profile the spatial covariance relationships that dictate protein folding arising from sequence variants in the population. Covariance analysis suggests a role for the ATPase activity of GRP94 in controlling the N- to C-terminal cooperative folding of alpha-1-antitrypsin responsible for the correction of liver aggregation and lung-disease phenotypes of alpha-1-antitrypsin deficiency. Gaussian process-based spatial covariance profiling provides a standard model built on covariant principles to evaluate the role of proteostasis components in guiding information flow from genome to proteome in response to genetic variation, potentially allowing us to intervene in the onset and progression of complex multi-system human diseases.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Capturing the conversion of the pathogenic alpha-1-antitrypsin fold by ATF6 enhanced proteostasis.Cell Chem Biol. 2023 Jan 19;30(1):22-42.e5. doi: 10.1016/j.chembiol.2022.12.004. Epub 2023 Jan 10. Cell Chem Biol. 2023. PMID: 36630963 Free PMC article.
-
Biological and structural basis for Aha1 regulation of Hsp90 ATPase activity in maintaining proteostasis in the human disease cystic fibrosis.Mol Biol Cell. 2010 Mar 15;21(6):871-84. doi: 10.1091/mbc.e09-12-1017. Epub 2010 Jan 20. Mol Biol Cell. 2010. PMID: 20089831 Free PMC article.
-
Interplay between the Hsp90 Chaperone and the HslVU Protease To Regulate the Level of an Essential Protein in Shewanella oneidensis.mBio. 2019 May 14;10(3):e00269-19. doi: 10.1128/mBio.00269-19. mBio. 2019. PMID: 31088919 Free PMC article.
-
The HSP90 chaperone machinery.Nat Rev Mol Cell Biol. 2017 Jun;18(6):345-360. doi: 10.1038/nrm.2017.20. Epub 2017 Apr 21. Nat Rev Mol Cell Biol. 2017. PMID: 28429788 Review.
-
Alpha 1-Antitrypsin Deficiency: A Disorder of Proteostasis-Mediated Protein Folding and Trafficking Pathways.Int J Mol Sci. 2020 Feb 21;21(4):1493. doi: 10.3390/ijms21041493. Int J Mol Sci. 2020. PMID: 32098273 Free PMC article. Review.
Cited by
-
The structural and functional dynamics of BiP and Grp94: opportunities for therapeutic discovery.Trends Pharmacol Sci. 2025 May;46(5):453-467. doi: 10.1016/j.tips.2025.03.004. Epub 2025 Apr 18. Trends Pharmacol Sci. 2025. PMID: 40253284 Review.
-
Spatial covariance reveals isothiocyanate natural products adjust redox stress to restore function in alpha-1-antitrypsin deficiency.Cell Rep Med. 2025 Jan 21;6(1):101917. doi: 10.1016/j.xcrm.2024.101917. Epub 2025 Jan 13. Cell Rep Med. 2025. PMID: 39809267 Free PMC article.
-
Hsp90: Bringing it all together.Cell Stress Chaperones. 2025 Feb;30(1):69-79. doi: 10.1016/j.cstres.2025.01.002. Epub 2025 Jan 29. Cell Stress Chaperones. 2025. PMID: 39889818 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
