

Generation of human alveolar epithelial type I cells from pluripotent stem cells
- PMID: 38642558
- PMCID: PMC11147407
- DOI: 10.1016/j.stem.2024.03.017
Generation of human alveolar epithelial type I cells from pluripotent stem cells
Abstract
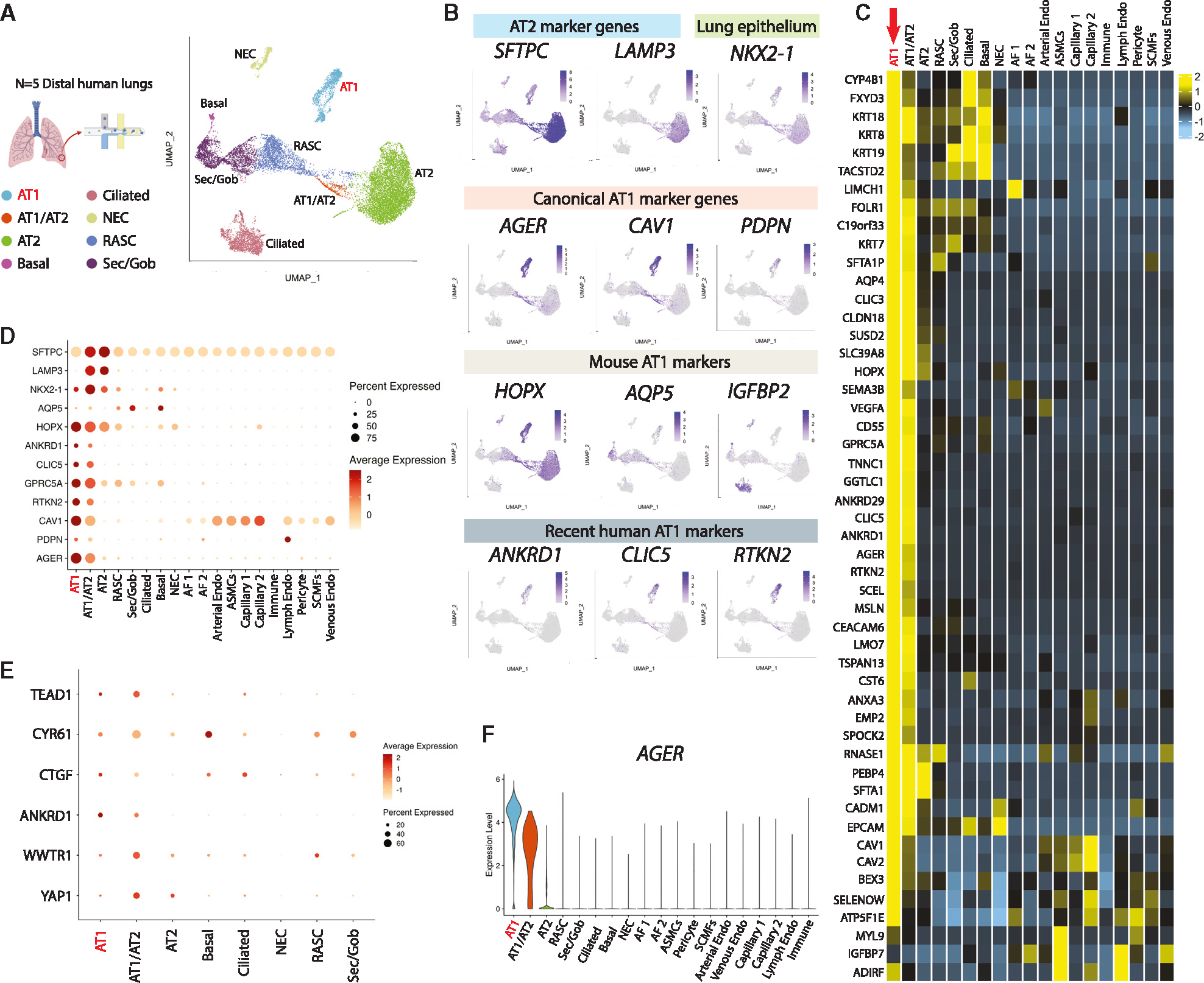
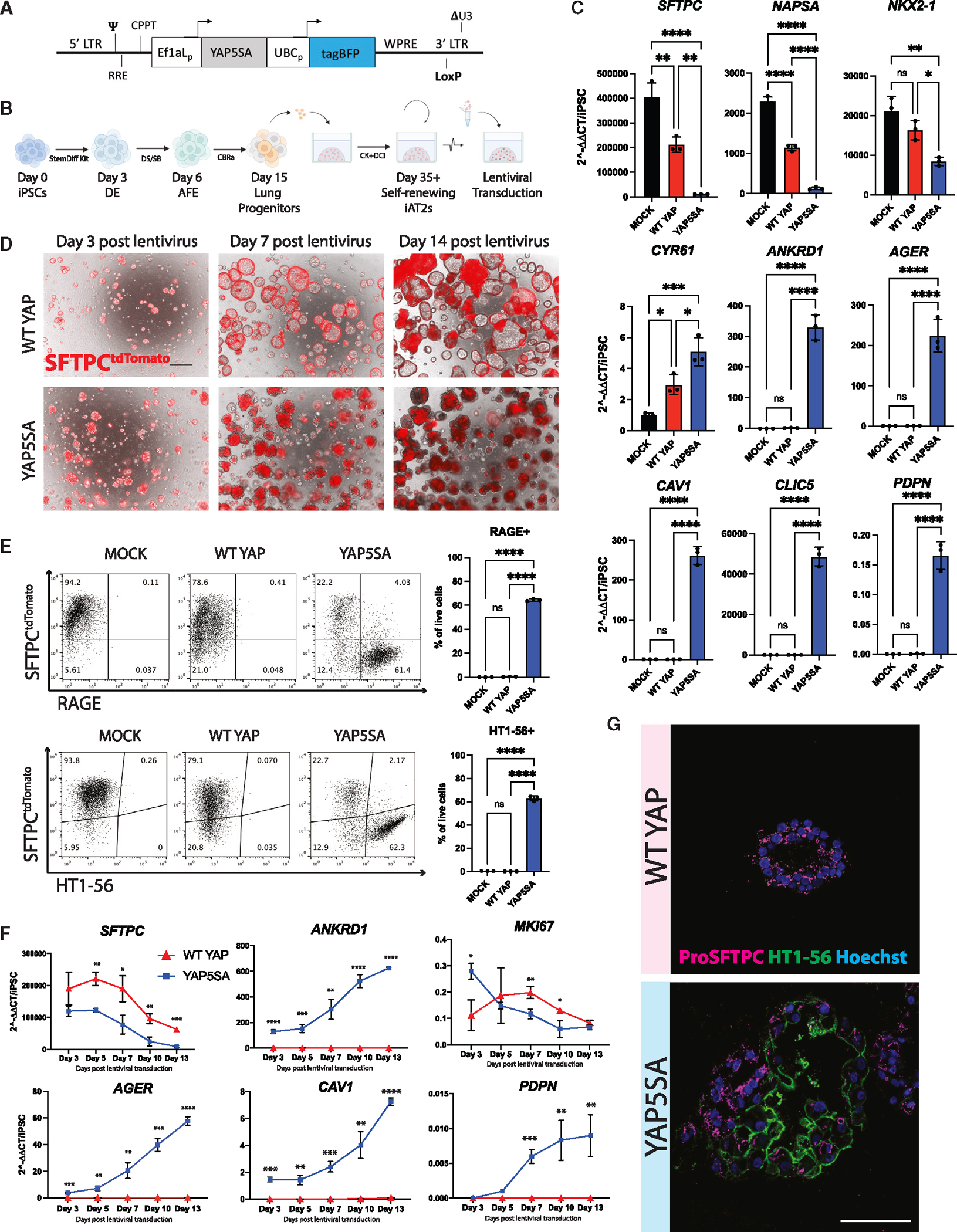
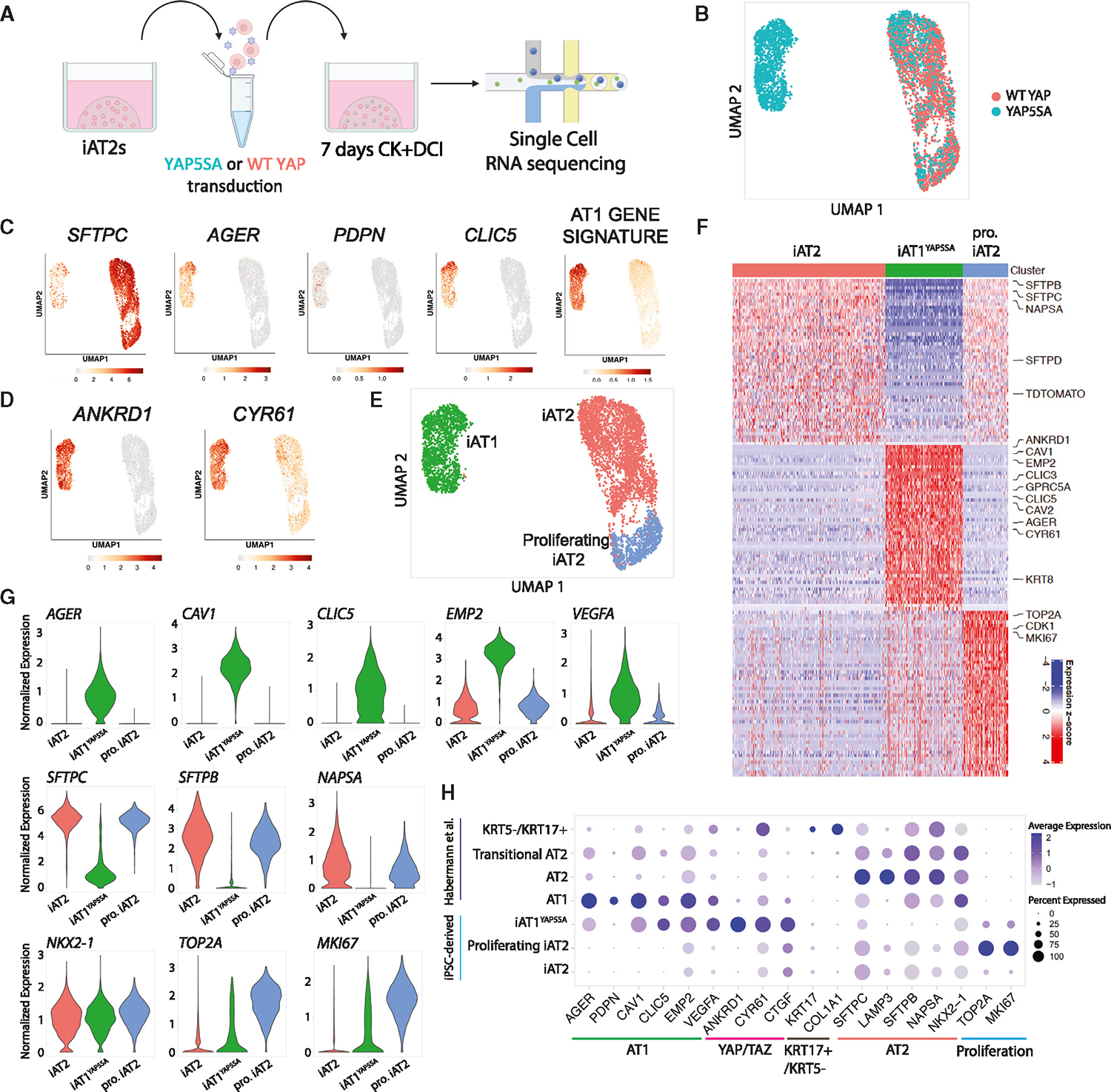
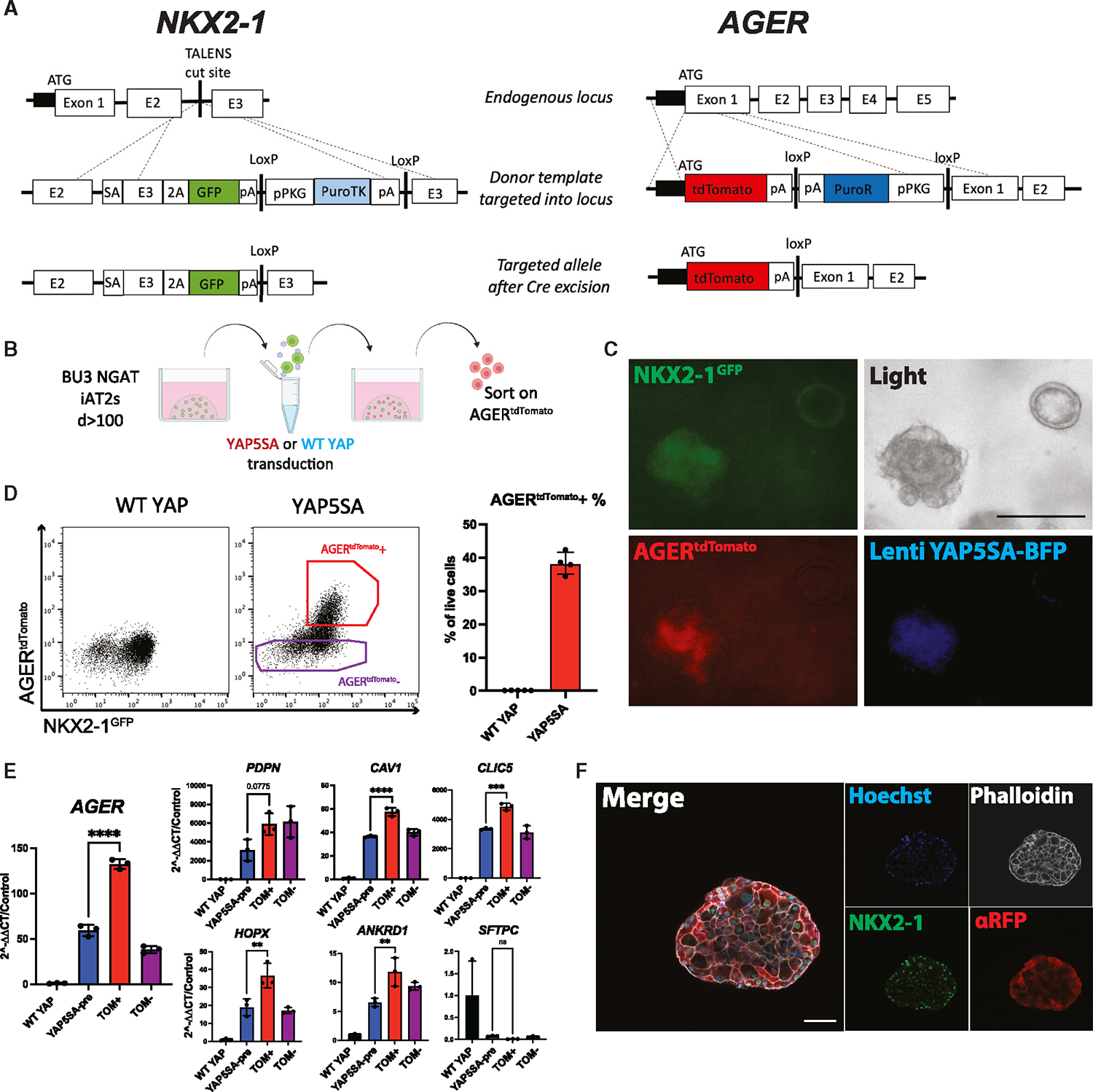
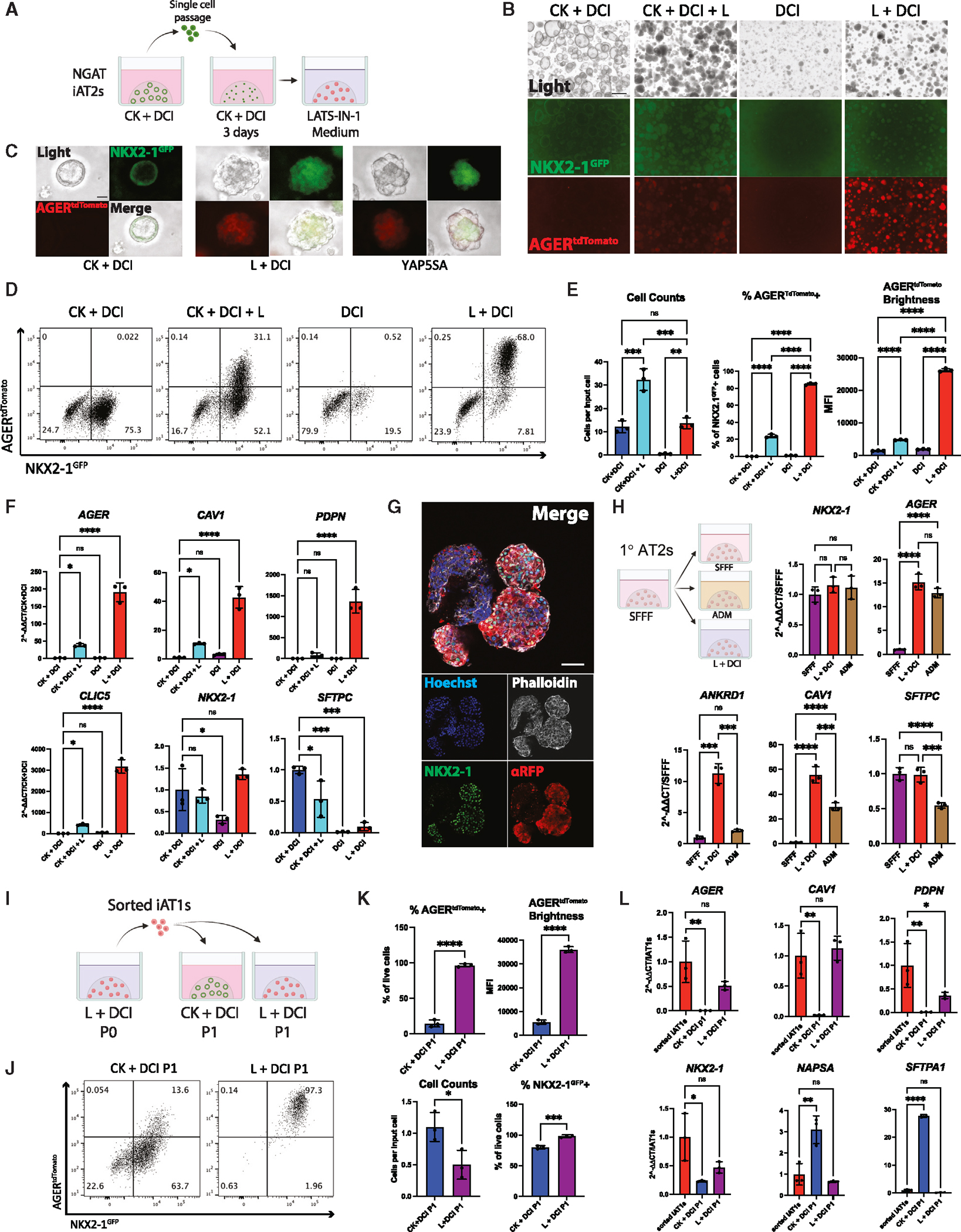
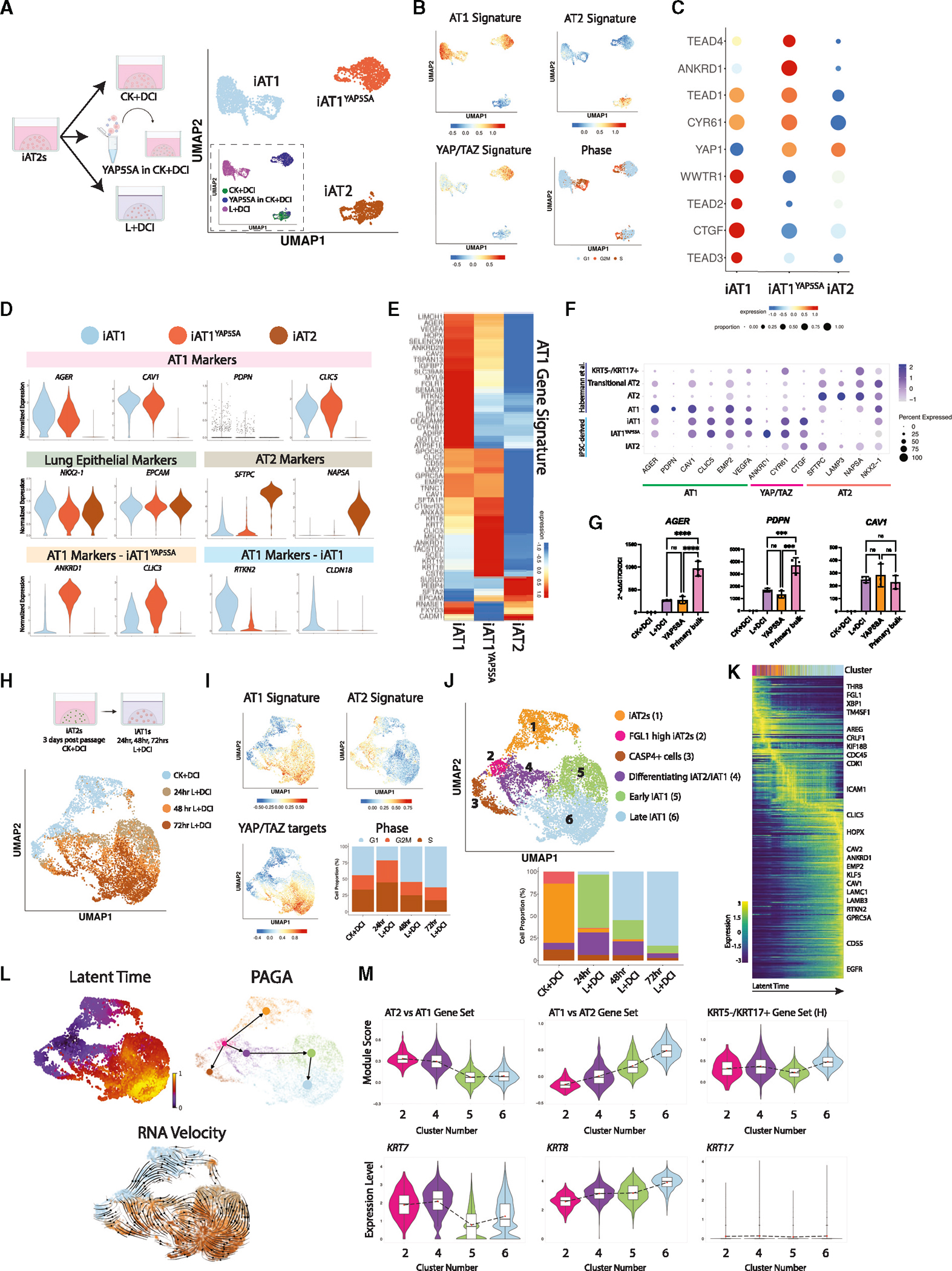
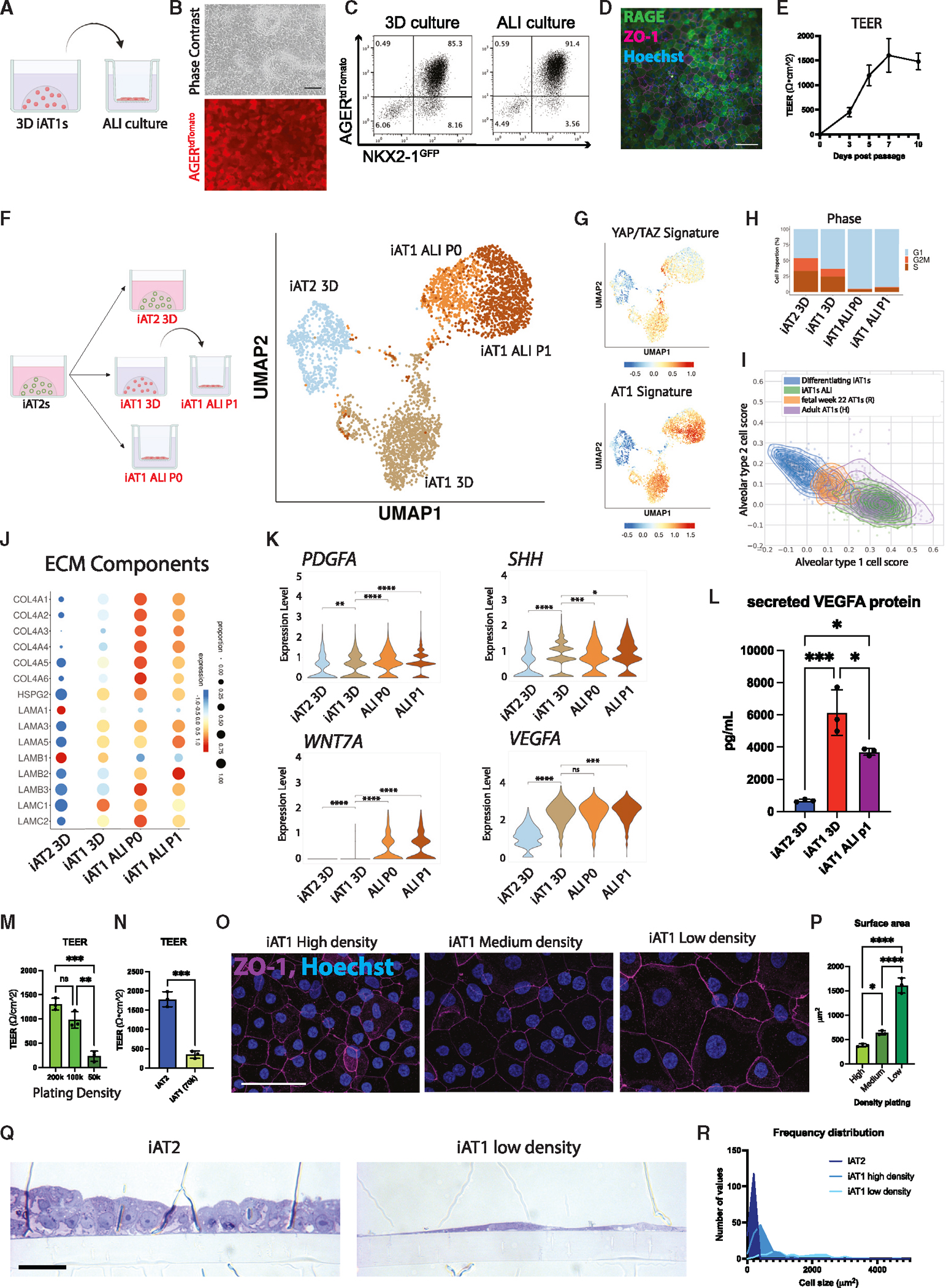
Alveolar epithelial type I cells (AT1s) line the gas exchange barrier of the distal lung and have been historically challenging to isolate or maintain in cell culture. Here, we engineer a human in vitro AT1 model system via directed differentiation of induced pluripotent stem cells (iPSCs). We use primary adult AT1 global transcriptomes to suggest benchmarks and pathways, such as Hippo-LATS-YAP/TAZ signaling, enriched in these cells. Next, we generate iPSC-derived alveolar epithelial type II cells (AT2s) and find that nuclear YAP signaling is sufficient to promote a broad transcriptomic shift from AT2 to AT1 gene programs. The resulting cells express a molecular, morphologic, and functional phenotype reminiscent of human AT1 cells, including the capacity to form a flat epithelial barrier producing characteristic extracellular matrix molecules and secreted ligands. Our results provide an in vitro model of human alveolar epithelial differentiation and a potential source of human AT1s.
Keywords: Hippo signaling; LATS inhibition; alveolar epithelial type I cells; directed differentiation; lung; lung epithelial reporter; pluripotent stem cells.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors have filed a patent application related to the generation of human alveolar epithelial type I cells from pluripotent stem cells detailed in this manuscript.
Figures
Update of
-
Generation of human alveolar epithelial type I cells from pluripotent stem cells.bioRxiv [Preprint]. 2023 Jan 20:2023.01.19.524655. doi: 10.1101/2023.01.19.524655. bioRxiv. 2023. Update in: Cell Stem Cell. 2024 May 2;31(5):657-675.e8. doi: 10.1016/j.stem.2024.03.017. PMID: 36711505 Free PMC article. Updated. Preprint.
References
-
- Habermann AC, Gutierrez AJ, Bui LT, Yahn SL, Winters NI, Calvi CL, Peter L, Chung MI, Taylor CJ, Jetter C, et al. (2020). Single-cell RNA sequencing reveals profibrotic roles of distinct epithelial and mesenchymal lineages in pulmonary fibrosis. Sci. Adv. 6, eaba1972. 10.1126/sciadv.aba1972. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
- U01 TR001810/TR/NCATS NIH HHS/United States
- R35 GM119461/GM/NIGMS NIH HHS/United States
- U01 HL152976/HL/NHLBI NIH HHS/United States
- K08 HL163494/HL/NHLBI NIH HHS/United States
- T32 HL007035/HL/NHLBI NIH HHS/United States
- U01 HL134745/HL/NHLBI NIH HHS/United States
- P30 ES013508/ES/NIEHS NIH HHS/United States
- R01 HL095993/HL/NHLBI NIH HHS/United States
- F31 HL158193/HL/NHLBI NIH HHS/United States
- R01 HL124392/HL/NHLBI NIH HHS/United States
- R01 HL164929/HL/NHLBI NIH HHS/United States
- U01 HL134766/HL/NHLBI NIH HHS/United States
- 75N92020C00005/HL/NHLBI NIH HHS/United States
- UL1 TR001430/TR/NCATS NIH HHS/United States
- P01 HL170952/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
