

Adipose-derived stem cells promote glycolysis and peritoneal metastasis via TGF-β1/SMAD3/ANGPTL4 axis in colorectal cancer
- PMID: 38643448
- PMCID: PMC11033247
- DOI: 10.1007/s00018-024-05215-1
Adipose-derived stem cells promote glycolysis and peritoneal metastasis via TGF-β1/SMAD3/ANGPTL4 axis in colorectal cancer
Abstract
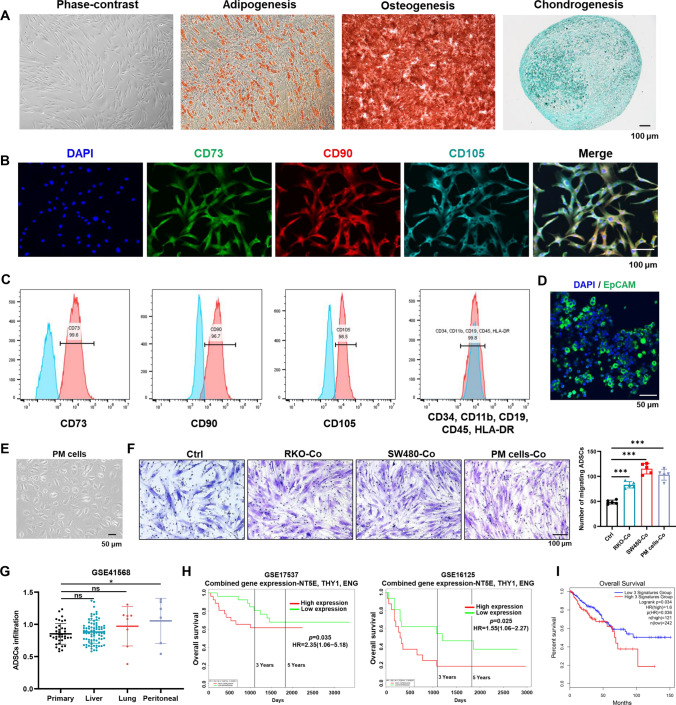
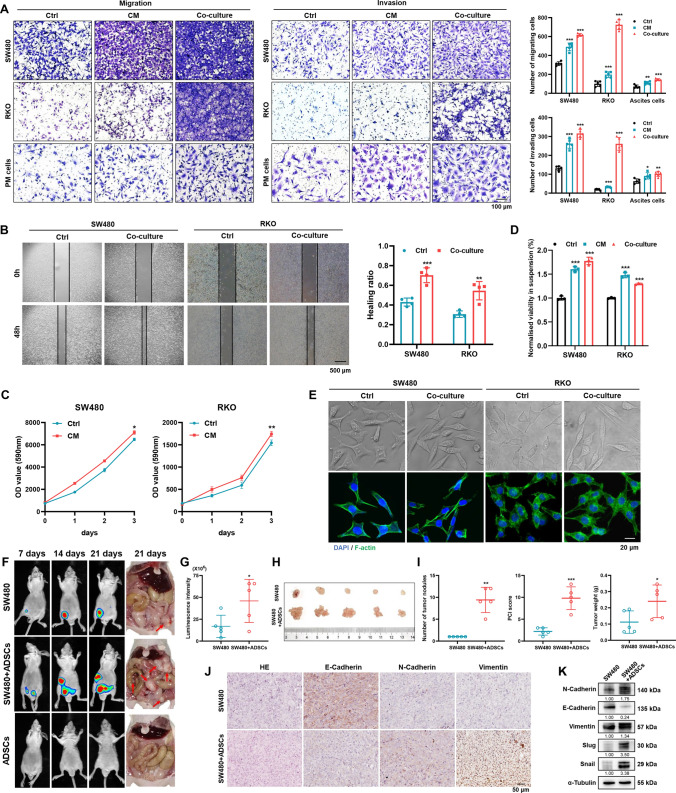
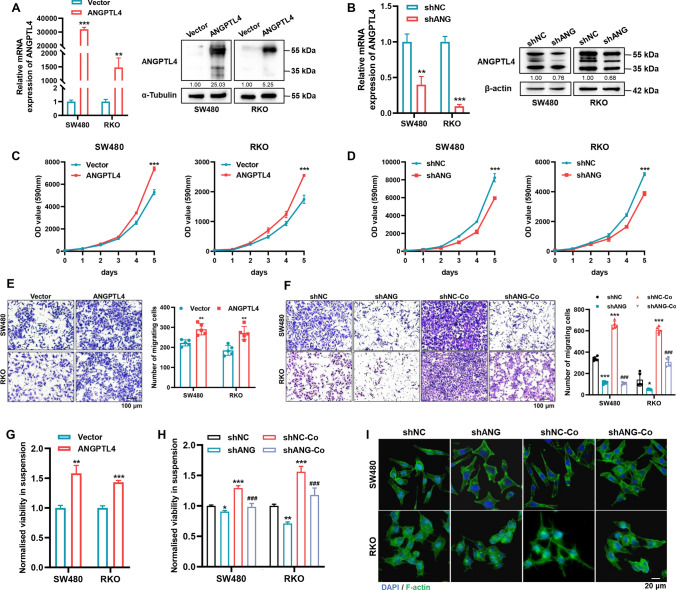
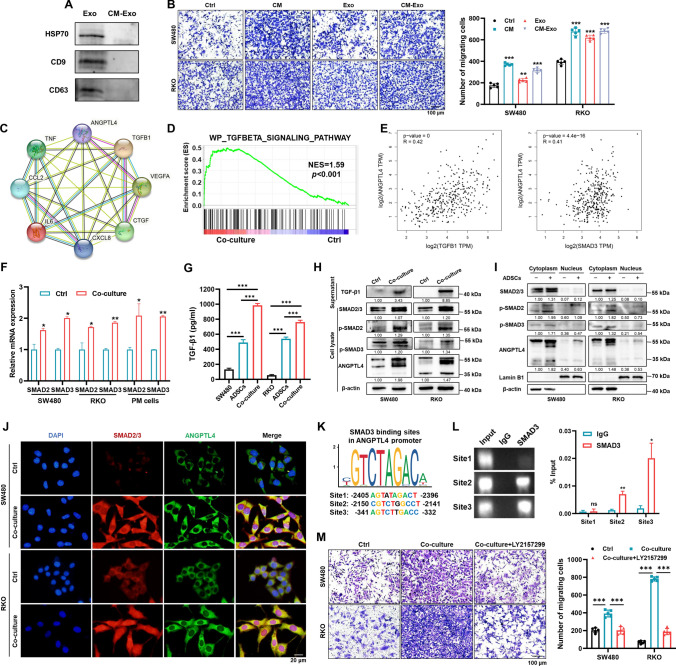
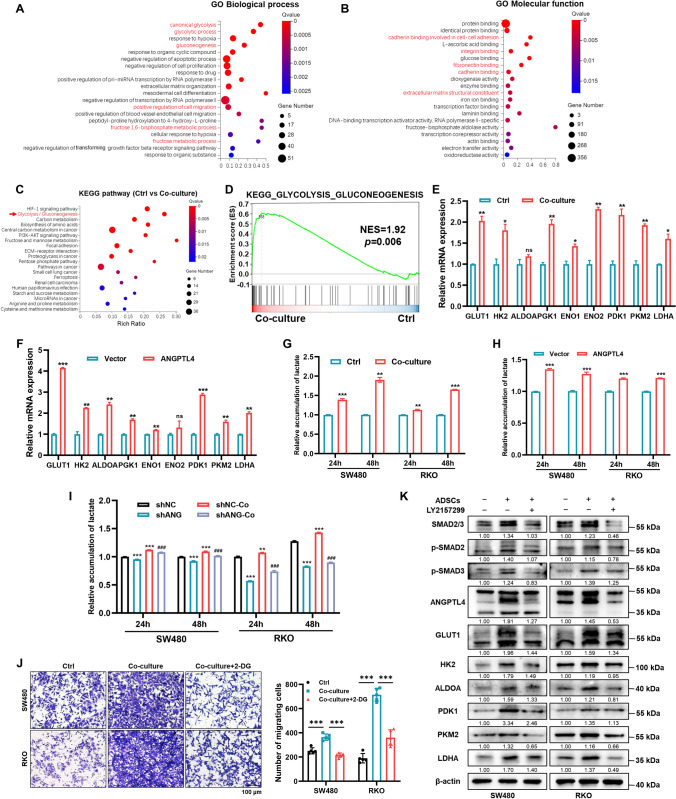
Peritoneal metastasis, the third most common metastasis in colorectal cancer (CRC), has a poor prognosis for the rapid progression and limited therapeutic strategy. However, the molecular characteristics and pathogenesis of CRC peritoneal metastasis are poorly understood. Here, we aimed to elucidate the action and mechanism of adipose-derived stem cells (ADSCs), a prominent component of the peritoneal microenvironment, in CRC peritoneal metastasis formation. Database analysis indicated that ADSCs infiltration was increased in CRC peritoneal metastases, and high expression levels of ADSCs marker genes predicted a poor prognosis. Then we investigated the effect of ADSCs on CRC cells in vitro and in vivo. The results revealed that CRC cells co-cultured with ADSCs exhibited stronger metastatic property and anoikis resistance, and ADSCs boosted the intraperitoneal seeding of CRC cells. Furthermore, RNA sequencing was carried out to identify the key target gene, angiopoietin like 4 (ANGPTL4), which was upregulated in CRC specimens, especially in peritoneal metastases. Mechanistically, TGF-β1 secreted by ADSCs activated SMAD3 in CRC cells, and chromatin immunoprecipitation assay showed that SMAD3 facilitated ANGPTL4 transcription by directly binding to ANGPTL4 promoter. The ANGPTL4 upregulation was essential for ADSCs to promote glycolysis and anoikis resistance in CRC. Importantly, simultaneously targeting TGF-β signaling and ANGPTL4 efficiently reduced intraperitoneal seeding in vivo. In conclusion, this study indicates that tumor-infiltrating ADSCs promote glycolysis and anoikis resistance in CRC cells and ultimately facilitate peritoneal metastasis via the TGF-β1/SMAD3/ANGPTL4 axis. The dual-targeting of TGF-β signaling and ANGPTL4 may be a feasible therapeutic strategy for CRC peritoneal metastasis.
Keywords: ANGPTL4; Adipose-derived mesenchymal stem cells; Colorectal carcinoma; Peritoneal carcinomatosis; TGF-β/SMAD; Warburg effect.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
MeSH terms
Substances
Grants and funding
- 82303366/National Natural Science Foundation of China
- 82203581/National Natural Science Foundation of China
- 2022M721526/Postdoctoral Research Foundation of China
- 2021A1515111220/Basic and Applied Basic Research Foundation of Guangdong Province
- 2022A1515110165/Basic and Applied Basic Research Foundation of Guangdong Province
LinkOut - more resources
Full Text Sources
Medical
