

This is a preprint.
Non-Invasive Label-free Analysis Pipeline for In Situ Characterization of Differentiation in 3D Brain Organoid Models
- PMID: 38645145
- PMCID: PMC11030508
- DOI: 10.21203/rs.3.rs-4049577/v1
Non-Invasive Label-free Analysis Pipeline for In Situ Characterization of Differentiation in 3D Brain Organoid Models
Update in
-
Non-invasive label-free imaging analysis pipeline for in situ characterization of 3D brain organoids.Sci Rep. 2024 Sep 27;14(1):22331. doi: 10.1038/s41598-024-72038-2. Sci Rep. 2024. PMID: 39333572 Free PMC article.
Abstract
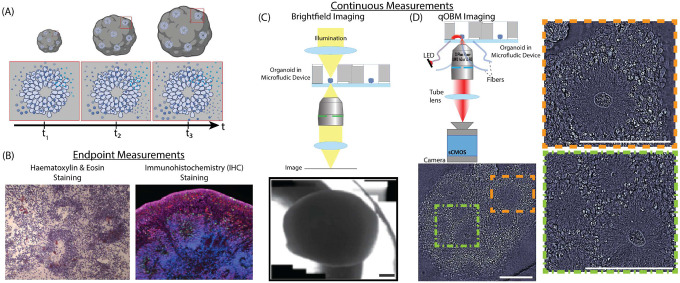
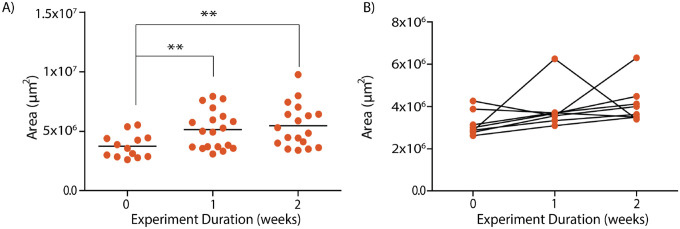
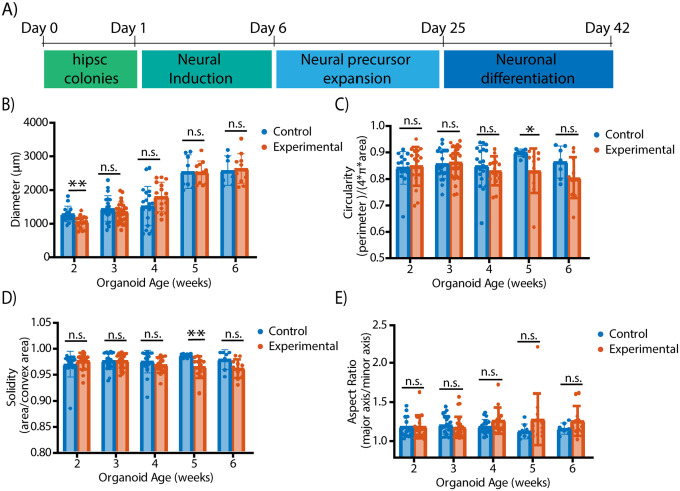
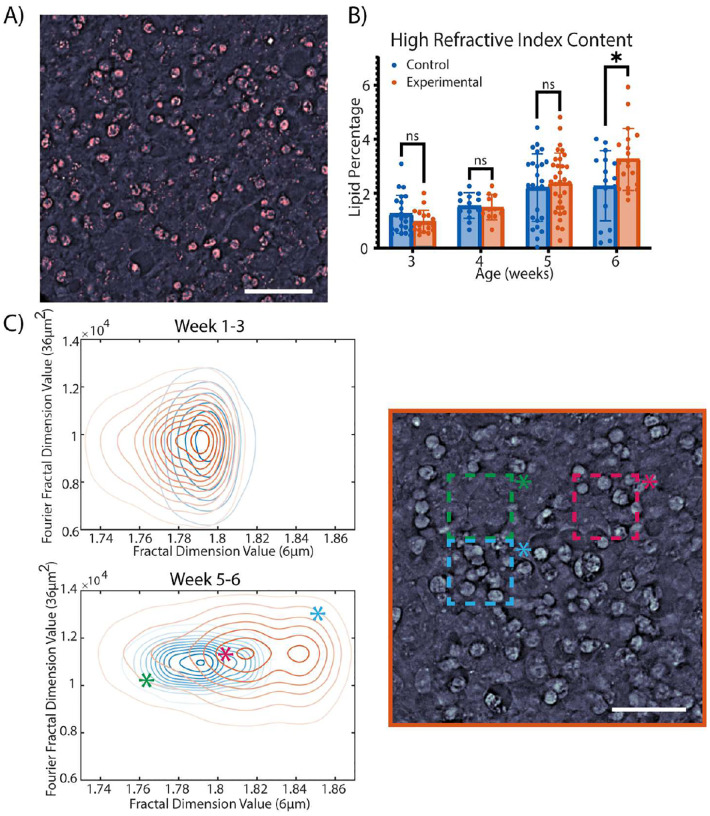
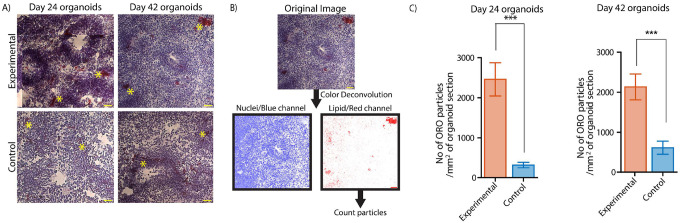
Brain organoids provide a unique opportunity to model organ development in a system similar to human organogenesis in vivo. Brain organoids thus hold great promise for drug screening and disease modeling. Conventional approaches to organoid characterization predominantly rely on molecular analysis methods, which are expensive, time-consuming, labor-intensive, and involve the destruction of the valuable 3D architecture of the organoids. This reliance on end-point assays makes it challenging to assess cellular and subcellular events occurring during organoid development in their 3D context. As a result, the long developmental processes are not monitored nor assessed. The ability to perform non-invasive assays is critical for longitudinally assessing features of organoid development during culture. In this paper, we demonstrate a label-free high-content imaging approach for observing changes in organoid morphology and structural changes occurring at the cellular and subcellular level. Enabled by microfluidic-based culture of 3D cell systems and a novel 3D quantitative phase imaging method, we demonstrate the ability to perform non-destructive high-resolution imaging of the organoid. The highlighted results demonstrated in this paper provide a new approach to performing live, non-destructive monitoring of organoid systems during culture.
Conflict of interest statement
Additional Declarations: No competing interests reported. The authors declare no competing interests
Figures


Similar articles
-
Non-invasive label-free imaging analysis pipeline for in situ characterization of 3D brain organoids.Sci Rep. 2024 Sep 27;14(1):22331. doi: 10.1038/s41598-024-72038-2. Sci Rep. 2024. PMID: 39333572 Free PMC article.
-
Development of a miniaturized 3D organoid culture platform for ultra-high-throughput screening.J Mol Cell Biol. 2020 Aug 1;12(8):630-643. doi: 10.1093/jmcb/mjaa036. J Mol Cell Biol. 2020. PMID: 32678871 Free PMC article.
-
Probing prodrug metabolism and reciprocal toxicity with an integrated and humanized multi-tissue organ-on-a-chip platform.Acta Biomater. 2020 Apr 1;106:124-135. doi: 10.1016/j.actbio.2020.02.015. Epub 2020 Feb 14. Acta Biomater. 2020. PMID: 32068138 Free PMC article.
-
Advances in Microfluidic Technologies in Organoid Research.Adv Healthc Mater. 2024 Aug;13(21):e2302686. doi: 10.1002/adhm.202302686. Epub 2023 Dec 28. Adv Healthc Mater. 2024. PMID: 38134345 Review.
-
Disease Modeling Using 3D Organoids Derived from Human Induced Pluripotent Stem Cells.Int J Mol Sci. 2018 Mar 21;19(4):936. doi: 10.3390/ijms19040936. Int J Mol Sci. 2018. PMID: 29561796 Free PMC article. Review.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
