

This is a preprint.
STING promotes homeostatic maintenance of tissues and confers longevity with aging
- PMID: 38645182
- PMCID: PMC11030237
- DOI: 10.1101/2024.04.04.588107
STING promotes homeostatic maintenance of tissues and confers longevity with aging
Abstract
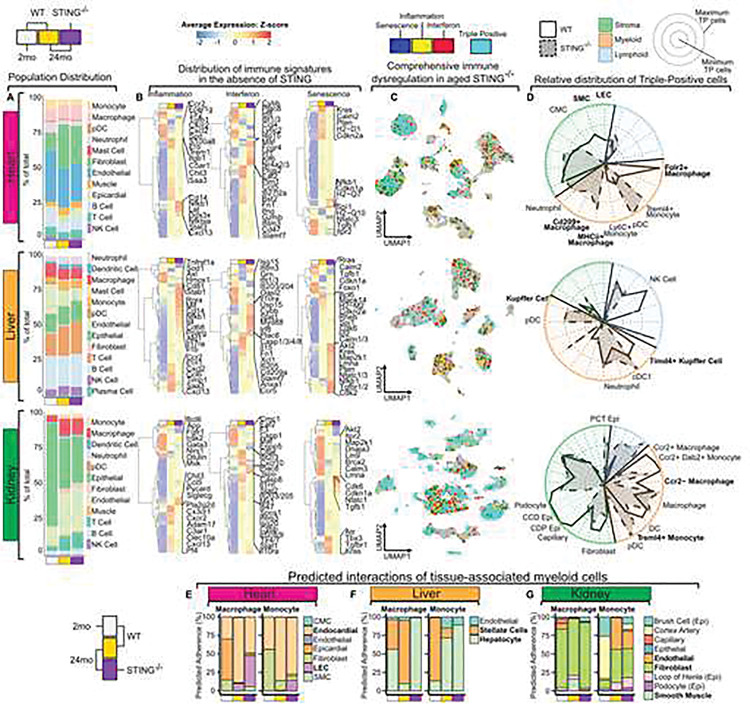
Local immune processes within aging tissues are a significant driver of aging associated dysfunction, but tissue-autonomous pathways and cell types that modulate these responses remain poorly characterized. The cytosolic DNA sensing pathway, acting through cyclic GMP-AMP synthase (cGAS) and Stimulator of Interferon Genes (STING), is broadly expressed in tissues, and is poised to regulate local type I interferon (IFN-I)-dependent and independent inflammatory processes within tissues. Recent studies suggest that the cGAS/STING pathway may drive pathology in various in vitro and in vivo models of accelerated aging. To date, however, the role of the cGAS/STING pathway in physiological aging processes, in the absence of genetic drivers, has remained unexplored. This remains a relevant gap, as STING is ubiquitously expressed, implicated in multitudinous disorders, and loss of function polymorphisms of STING are highly prevalent in the human population (>50%). Here we reveal that, during physiological aging, STING-deficiency leads to a significant shortening of murine lifespan, increased pro-inflammatory serum cytokines and tissue infiltrates, as well as salient changes in histological composition and organization. We note that aging hearts, livers, and kidneys express distinct subsets of inflammatory, interferon-stimulated gene (ISG), and senescence genes, collectively comprising an immune fingerprint for each tissue. These distinctive patterns are largely imprinted by tissue-specific stromal and myeloid cells. Using cellular interaction network analyses, immunofluorescence, and histopathology data, we show that these immune fingerprints shape the tissue architecture and the landscape of cell-cell interactions in aging tissues. These age-associated immune fingerprints are grossly dysregulated with STING-deficiency, with key genes that define aging STING-sufficient tissues greatly diminished in the absence of STING. Changes in immune signatures are concomitant with a restructuring of the stromal and myeloid fractions, whereby cell:cell interactions are grossly altered and resulting in disorganization of tissue architecture in STING-deficient organs. This altered homeostasis in aging STING-deficient tissues is associated with a cross-tissue loss of homeostatic tissue-resident macrophage (TRM) populations in these tissues. Ex vivo analyses reveal that basal STING-signaling limits the susceptibility of TRMs to death-inducing stimuli and determines their in situ localization in tissue niches, thereby promoting tissue homeostasis. Collectively, these data upend the paradigm that cGAS/STING signaling is primarily pathological in aging and instead indicate that basal STING signaling sustains tissue function and supports organismal longevity. Critically, our study urges caution in the indiscriminate targeting of these pathways, which may result in unpredictable and pathological consequences for health during aging.
Conflict of interest statement
DECLARATION OF INTERESTS The authors declare no competing interests.
Figures





References
-
- Lei Y., Guerra Martinez C., Torres-Odio S., Bell S.L., Birdwell C.E., Bryant J.D., Tong C.W., Watson R.O., West L.C., and West A.P. (2021). Elevated type I interferon responses potentiate metabolic dysfunction, inflammation, and accelerated aging in mtDNA mutator mice. Sci Adv 7. 10.1126/sciadv.abe7548. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous