

Extensive Loss and Gain of Conserved Noncoding Elements During Early Teleost Evolution
- PMID: 38648507
- PMCID: PMC11034925
- DOI: 10.1093/gbe/evae061
Extensive Loss and Gain of Conserved Noncoding Elements During Early Teleost Evolution
Abstract
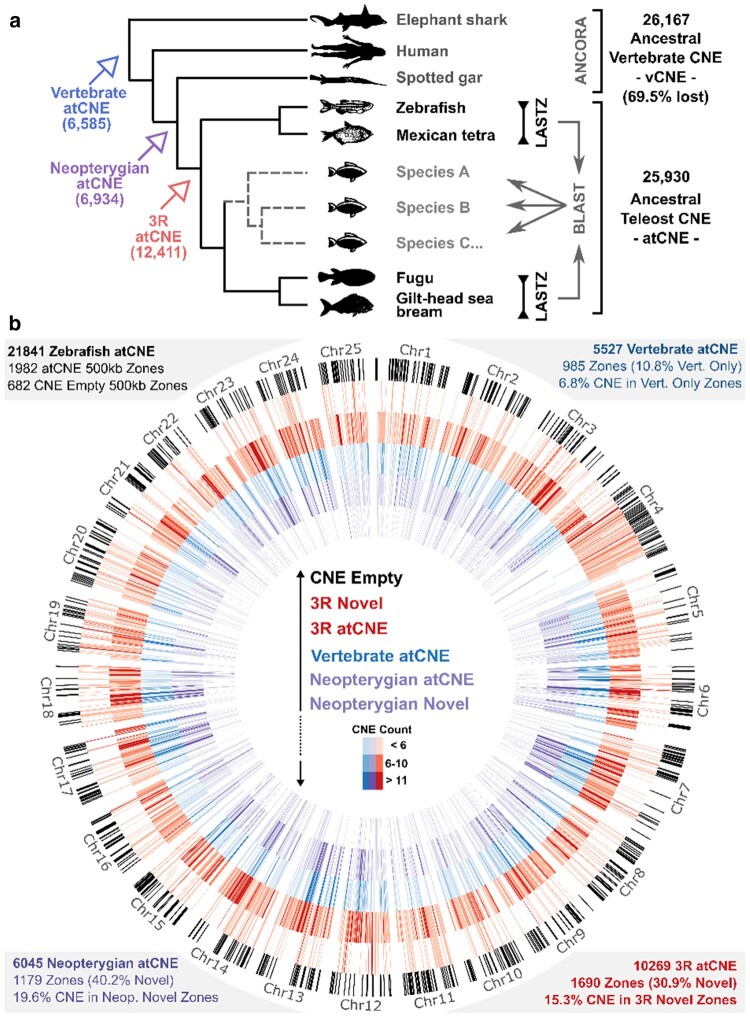
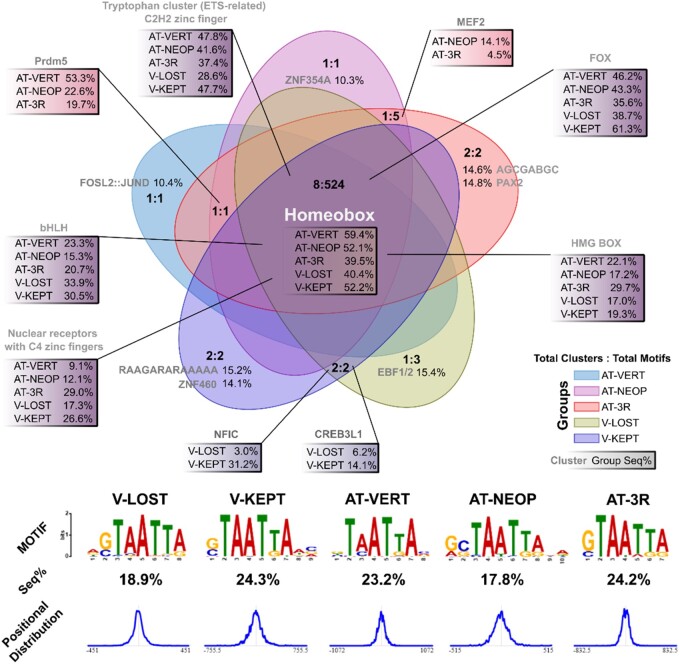
Conserved noncoding elements in vertebrates are enriched around transcription factor loci associated with development. However, loss and rapid divergence of conserved noncoding elements has been reported in teleost fish, albeit taking only few genomes into consideration. Taking advantage of the recent increase in high-quality teleost genomes, we focus on studying the evolution of teleost conserved noncoding elements, carrying out targeted genomic alignments and comparisons within the teleost phylogeny to detect conserved noncoding elements and reconstruct the ancestral teleost conserved noncoding elements repertoire. This teleost-centric approach confirms previous observations of extensive vertebrate conserved noncoding elements loss early in teleost evolution, but also reveals massive conserved noncoding elements gain in the teleost stem-group over 300 million years ago. Using synteny-based association to link conserved noncoding elements to their putatively regulated target genes, we show the most teleost gained conserved noncoding elements are found in the vicinity of orthologous loci involved in transcriptional regulation and embryonic development that are also associated with conserved noncoding elements in other vertebrates. Moreover, teleost and vertebrate conserved noncoding elements share a highly similar motif and transcription factor binding site vocabulary. We suggest that early teleost conserved noncoding element gains reflect a restructuring of the ancestral conserved noncoding element repertoire through both extreme divergence and de novo emergence. Finally, we support newly identified pan-teleost conserved noncoding elements have potential for accurate resolution of teleost phylogenetic placements in par with coding sequences, unlike ancestral only elements shared with spotted gar. This work provides new insight into conserved noncoding element evolution with great value for follow-up work on phylogenomics, comparative genomics, and the study of gene regulation evolution in teleosts.
Keywords: CNE; conserved noncoding elements; enhancer evolution; teleost comparative genomics; whole-genome duplication.
© The Author(s) 2024. Published by Oxford University Press on behalf of Society for Molecular Biology and Evolution.
Figures




References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
