

Biomolecular condensates form spatially inhomogeneous network fluids
- PMID: 38649740
- PMCID: PMC11035652
- DOI: 10.1038/s41467-024-47602-z
Biomolecular condensates form spatially inhomogeneous network fluids
Abstract
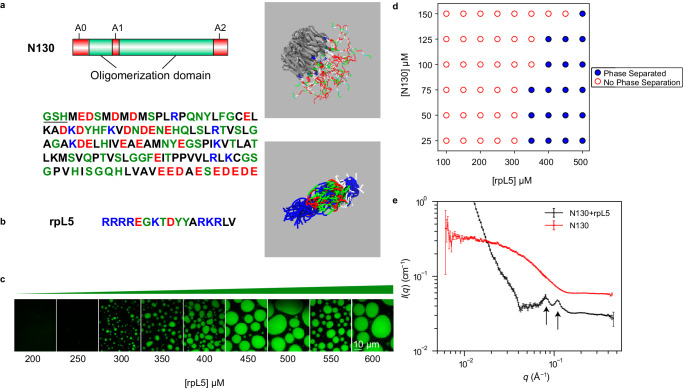
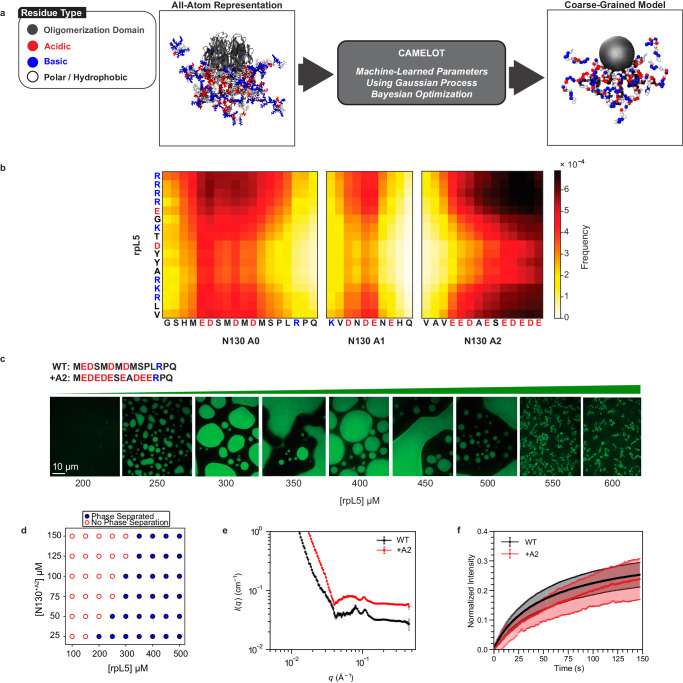
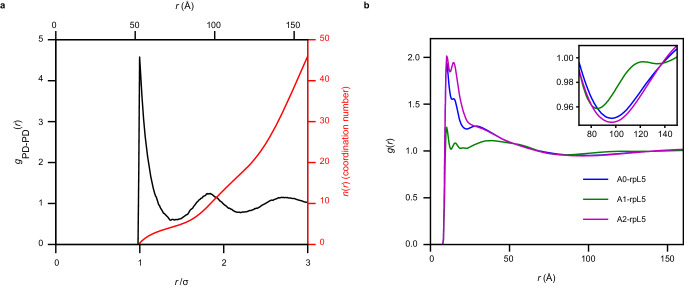
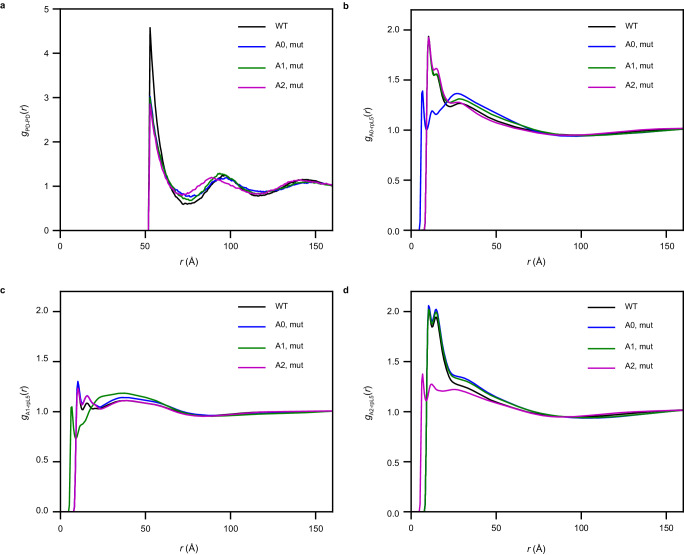
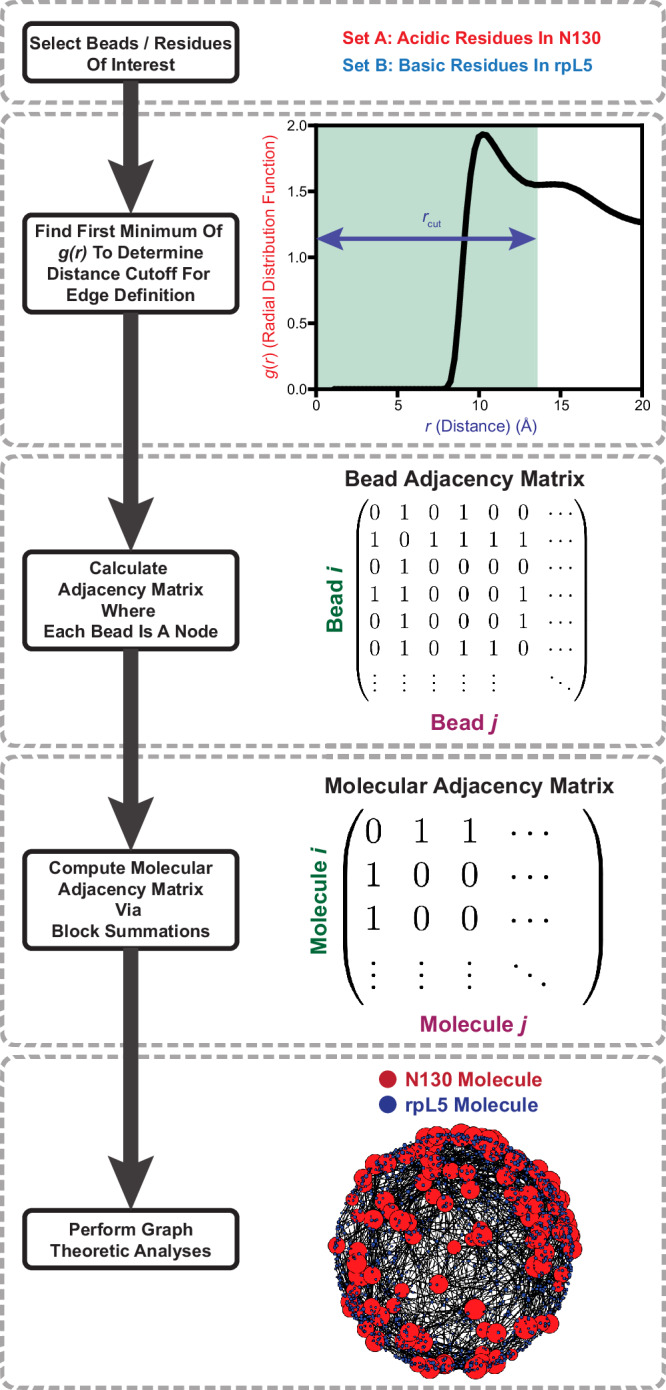
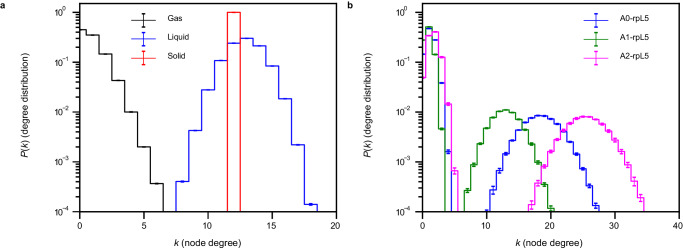
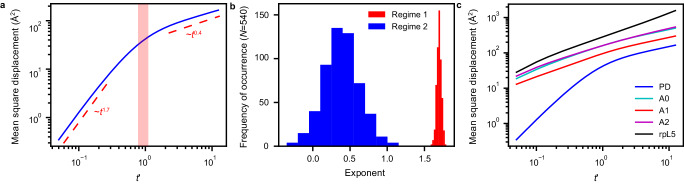
The functions of biomolecular condensates are thought to be influenced by their material properties, and these will be determined by the internal organization of molecules within condensates. However, structural characterizations of condensates are challenging, and rarely reported. Here, we deploy a combination of small angle neutron scattering, fluorescence recovery after photobleaching, and coarse-grained molecular dynamics simulations to provide structural descriptions of model condensates that are formed by macromolecules from nucleolar granular components (GCs). We show that these minimal facsimiles of GCs form condensates that are network fluids featuring spatial inhomogeneities across different length scales that reflect the contributions of distinct protein and peptide domains. The network-like inhomogeneous organization is characterized by a coexistence of liquid- and gas-like macromolecular densities that engenders bimodality of internal molecular dynamics. These insights suggest that condensates formed by multivalent proteins share features with network fluids formed by systems such as patchy or hairy colloids.
© 2024. The Author(s).
Conflict of interest statement
R.V.P. is a member of the scientific advisory board and shareholder of Dewpoint Therapeutics Inc. D.M.M. is an employee and shareholder of Dewpoint Therapeutics. The work reported here was not influenced by these affiliations. The remaining authors have no competing interests to declare.
Figures
Update of
-
Biomolecular condensates form spatially inhomogeneous network fluids.bioRxiv [Preprint]. 2024 Feb 11:2023.10.07.561338. doi: 10.1101/2023.10.07.561338. bioRxiv. 2024. Update in: Nat Commun. 2024 Apr 22;15(1):3413. doi: 10.1038/s41467-024-47602-z. PMID: 37873180 Free PMC article. Updated. Preprint.
-
Biomolecular condensates form spatially inhomogeneous network fluids.Res Sq [Preprint]. 2023 Oct 18:rs.3.rs-3419423. doi: 10.21203/rs.3.rs-3419423/v1. Res Sq. 2023. Update in: Nat Commun. 2024 Apr 22;15(1):3413. doi: 10.1038/s41467-024-47602-z. PMID: 37886520 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
