

PeNAC67-PeKAN2-PeSCL23 and B-class MADS-box transcription factors synergistically regulate the specialization process from petal to lip in Phalaenopsis equestris
- PMID: 38649966
- PMCID: PMC11036780
- DOI: 10.1186/s43897-023-00079-8
PeNAC67-PeKAN2-PeSCL23 and B-class MADS-box transcription factors synergistically regulate the specialization process from petal to lip in Phalaenopsis equestris
Abstract
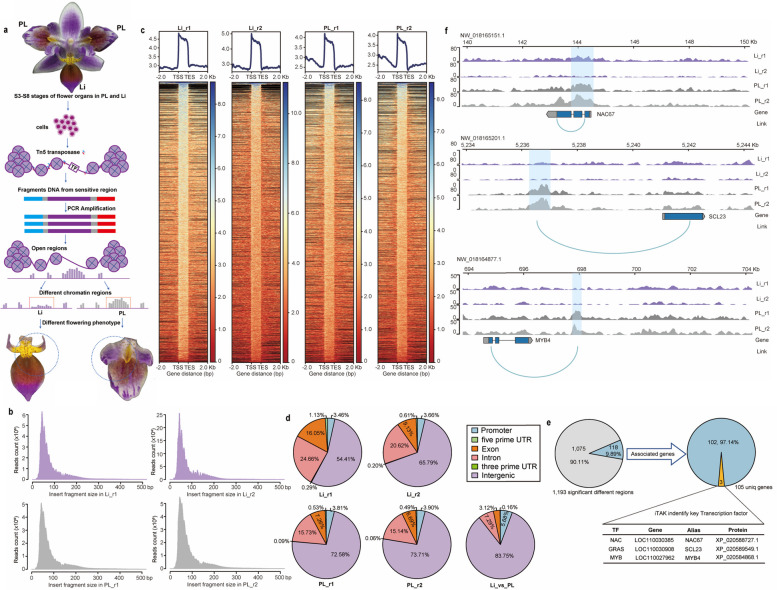
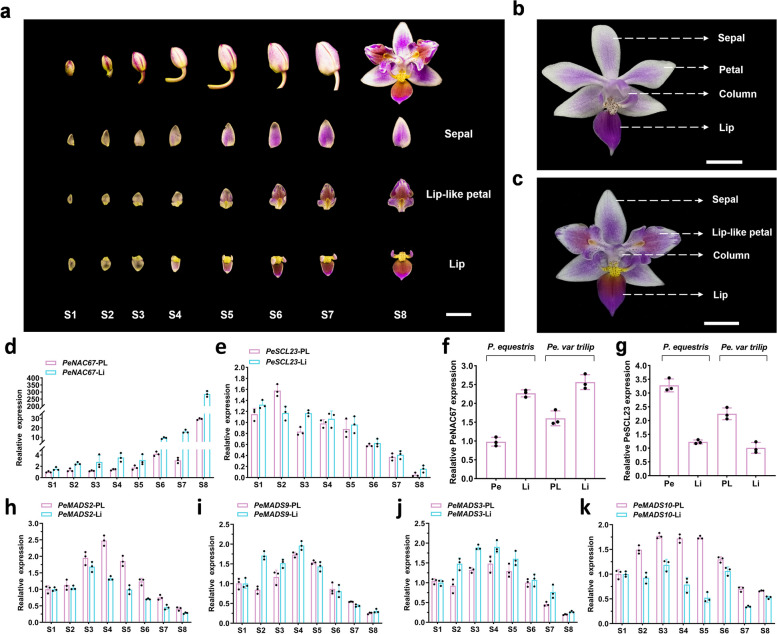
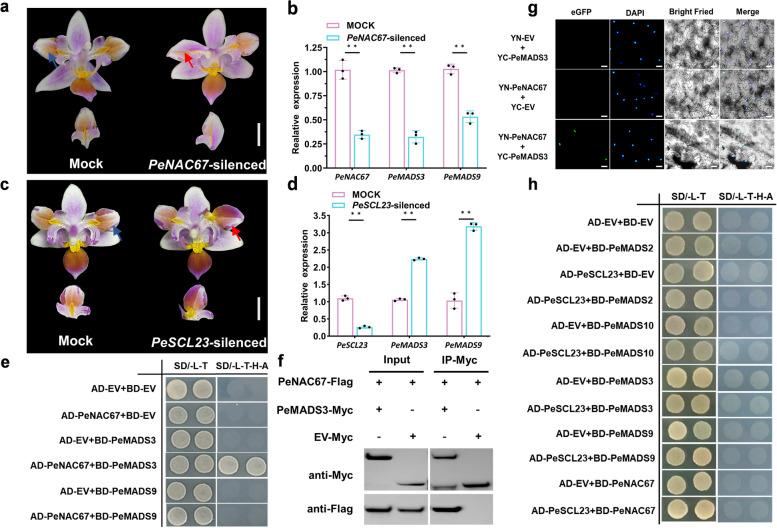
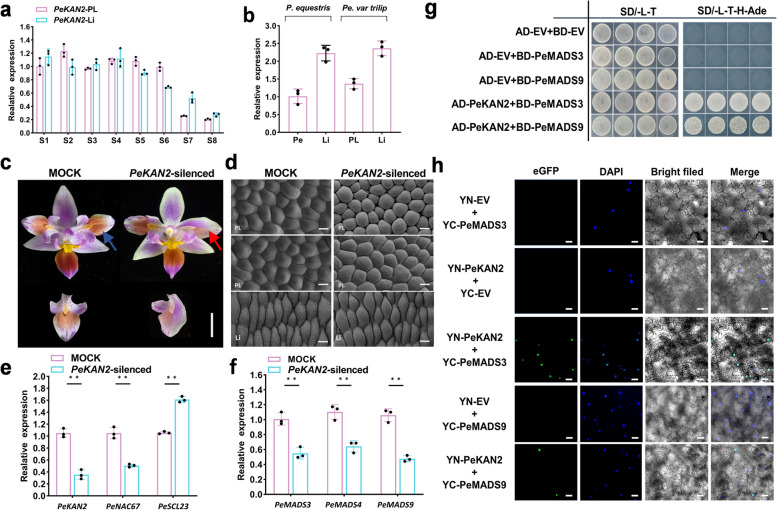
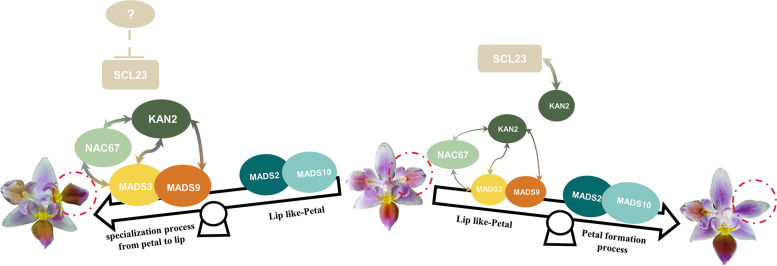
The molecular basis of orchid flower development involves a specific regulatory program in which MADS-box transcription factors play a central role. The recent 'perianth code' model hypothesizes that two types of higher-order heterotetrameric complexes, namely SP complex and L complex, play pivotal roles in the orchid perianth organ formation. Therefore, we explored their roles and searched for other components of the regulatory network.Through the combined analysis for transposase-accessible chromatin with high-throughput sequencing and RNA sequencing of the lip-like petal and lip from Phalaenopsis equestris var.trilip, transcription factor-(TF) genes involved in lip development were revealed. PeNAC67 encoding a NAC-type TF and PeSCL23 encoding a GRAS-type TF were differentially expressed between the lip-like petal and the lip. PeNAC67 interacted with and stabilized PeMADS3, which positively regulated the development of lip-like petal to lip. PeSCL23 and PeNAC67 competitively bound with PeKAN2 and positively regulated the development of lip-like petal to petal by affecting the level of PeMADS3. PeKAN2 as an important TF that interacts with PeMADS3 and PeMADS9 can promote lip development. These results extend the 'perianth code' model and shed light on the complex regulation of orchid flower development.
Keywords: PeKAN2; PeNAC67; PeSCL23; Phalaenopsis orchids; Lip development; ‘Perianth code’ model.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


Similar articles
-
Extending the Toolkit for Beauty: Differential Co-Expression of DROOPING LEAF-Like and Class B MADS-Box Genes during Phalaenopsis Flower Development.Int J Mol Sci. 2021 Jun 29;22(13):7025. doi: 10.3390/ijms22137025. Int J Mol Sci. 2021. PMID: 34209912 Free PMC article.
-
Virus-induced gene silencing unravels multiple transcription factors involved in floral growth and development in Phalaenopsis orchids.J Exp Bot. 2013 Sep;64(12):3869-84. doi: 10.1093/jxb/ert218. J Exp Bot. 2013. PMID: 23956416 Free PMC article.
-
A tetraspanin gene regulating auxin response and affecting orchid perianth size and various plant developmental processes.Plant Direct. 2019 Aug 5;3(8):e00157. doi: 10.1002/pld3.157. eCollection 2019 Aug. Plant Direct. 2019. PMID: 31406958 Free PMC article.
-
How to Evolve a Perianth: A Review of Cadastral Mechanisms for Perianth Identity.Front Plant Sci. 2018 Oct 29;9:1573. doi: 10.3389/fpls.2018.01573. eCollection 2018. Front Plant Sci. 2018. PMID: 30420867 Free PMC article. Review.
-
Why are orchid flowers so diverse? Reduction of evolutionary constraints by paralogues of class B floral homeotic genes.Ann Bot. 2009 Aug;104(3):583-94. doi: 10.1093/aob/mcn258. Epub 2009 Jan 13. Ann Bot. 2009. PMID: 19141602 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous