

A tryptophan-derived uremic metabolite/Ahr/Pdk4 axis governs skeletal muscle mitochondrial energetics in chronic kidney disease
- PMID: 38652558
- PMCID: PMC11141944
- DOI: 10.1172/jci.insight.178372
A tryptophan-derived uremic metabolite/Ahr/Pdk4 axis governs skeletal muscle mitochondrial energetics in chronic kidney disease
Abstract
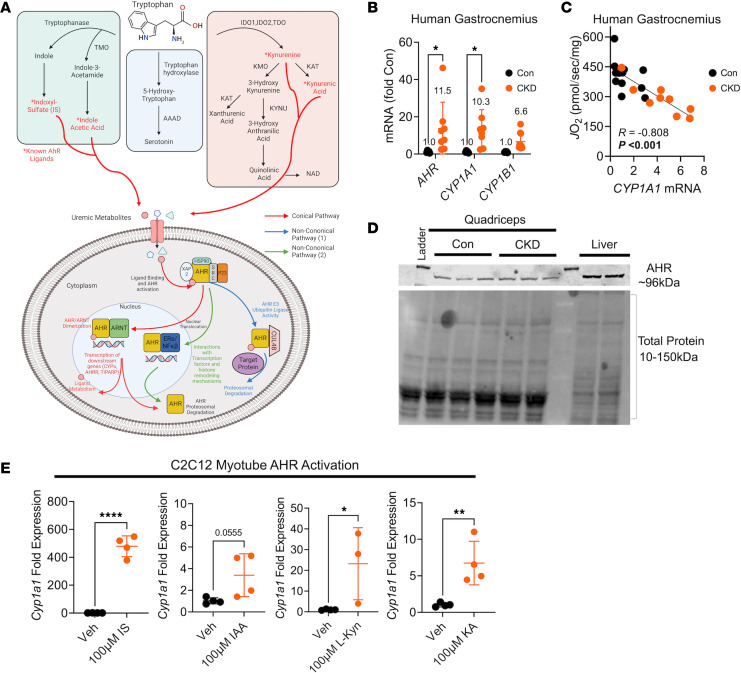
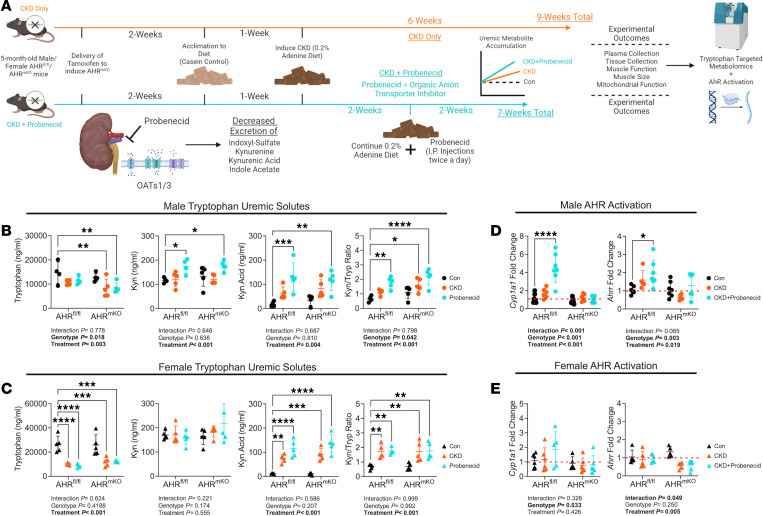
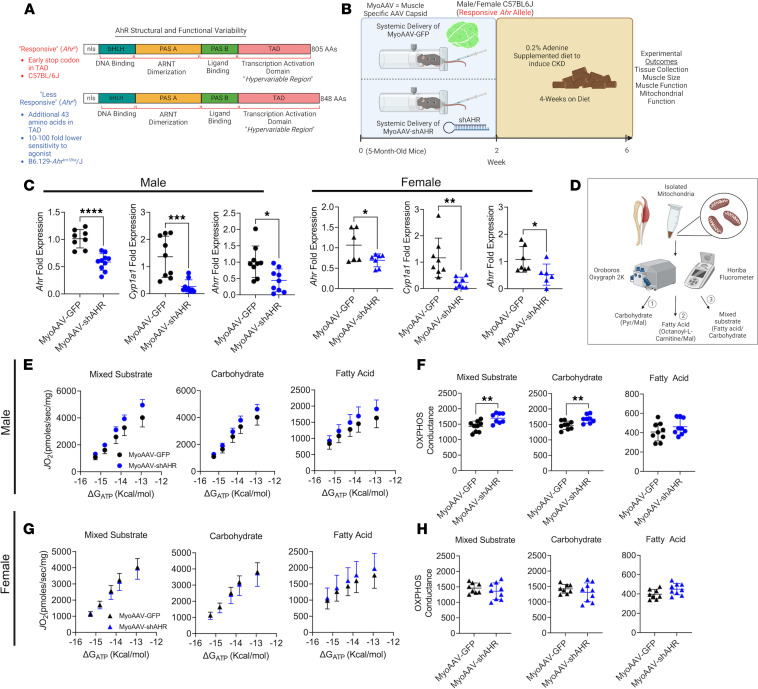
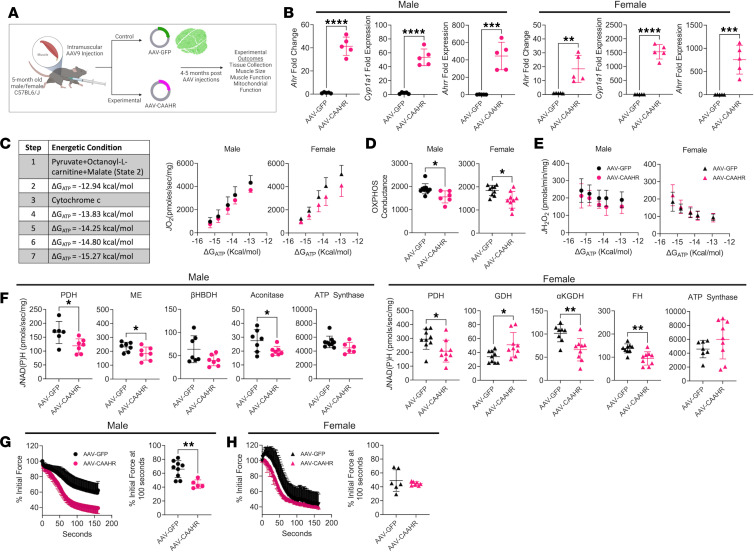
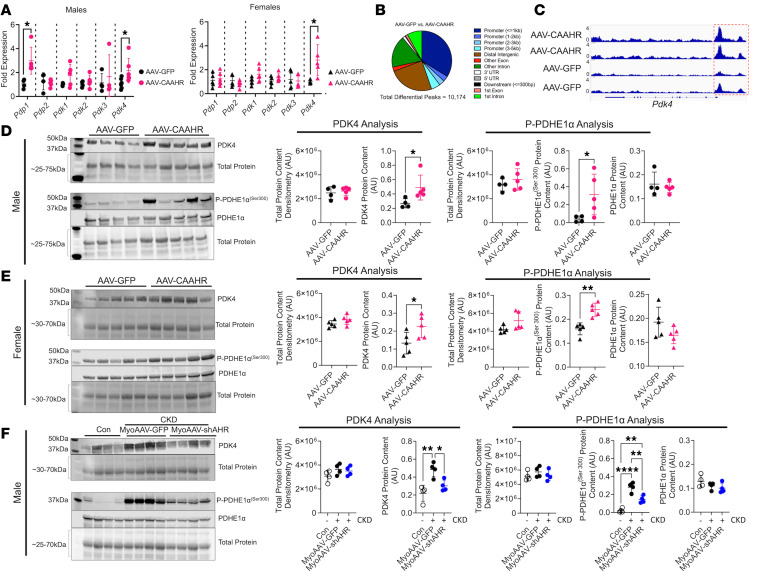
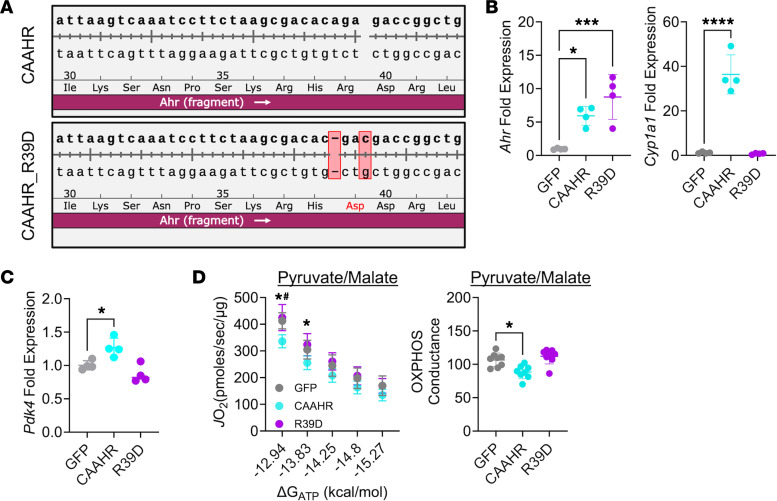
Chronic kidney disease (CKD) causes accumulation of uremic metabolites that negatively affect skeletal muscle. Tryptophan-derived uremic metabolites are agonists of the aryl hydrocarbon receptor (AHR), which has been shown to be activated in CKD. This study investigated the role of the AHR in skeletal muscle pathology of CKD. Compared with controls with normal kidney function, AHR-dependent gene expression (CYP1A1 and CYP1B1) was significantly upregulated in skeletal muscle of patients with CKD, and the magnitude of AHR activation was inversely correlated with mitochondrial respiration. In mice with CKD, muscle mitochondrial oxidative phosphorylation (OXPHOS) was markedly impaired and strongly correlated with the serum level of tryptophan-derived uremic metabolites and AHR activation. Muscle-specific deletion of the AHR substantially improved mitochondrial OXPHOS in male mice with the greatest uremic toxicity (CKD + probenecid) and abolished the relationship between uremic metabolites and OXPHOS. The uremic metabolite/AHR/mitochondrial axis in skeletal muscle was verified using muscle-specific AHR knockdown in C57BL/6J mice harboring a high-affinity AHR allele, as well as ectopic viral expression of constitutively active mutant AHR in mice with normal renal function. Notably, OXPHOS changes in AHRmKO mice were present only when mitochondria were fueled by carbohydrates. Further analyses revealed that AHR activation in mice led to significantly increased pyruvate dehydrogenase kinase 4 (Pdk4) expression and phosphorylation of pyruvate dehydrogenase enzyme. These findings establish a uremic metabolite/AHR/Pdk4 axis in skeletal muscle that governs mitochondrial deficits in carbohydrate oxidation during CKD.
Keywords: Chronic kidney disease; Mitochondria; Muscle biology; Nephrology; Skeletal muscle.
Conflict of interest statement
Figures

References
-
- Global Burden of Disease Study 2013 Collaborators Global, regional, and national incidence, prevalence, and years lived with disability for 301 acute and chronic diseases and injuries in 188 countries, 1990-2013: a systematic analysis for the Global Burden of Disease Study 2013. Lancet. 2015;386(9995):743–800. doi: 10.1016/S0140-6736(15)60692-4. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
