

DrugMap: A quantitative pan-cancer analysis of cysteine ligandability
- PMID: 38653237
- PMCID: PMC11143475
- DOI: 10.1016/j.cell.2024.03.027
DrugMap: A quantitative pan-cancer analysis of cysteine ligandability
Abstract
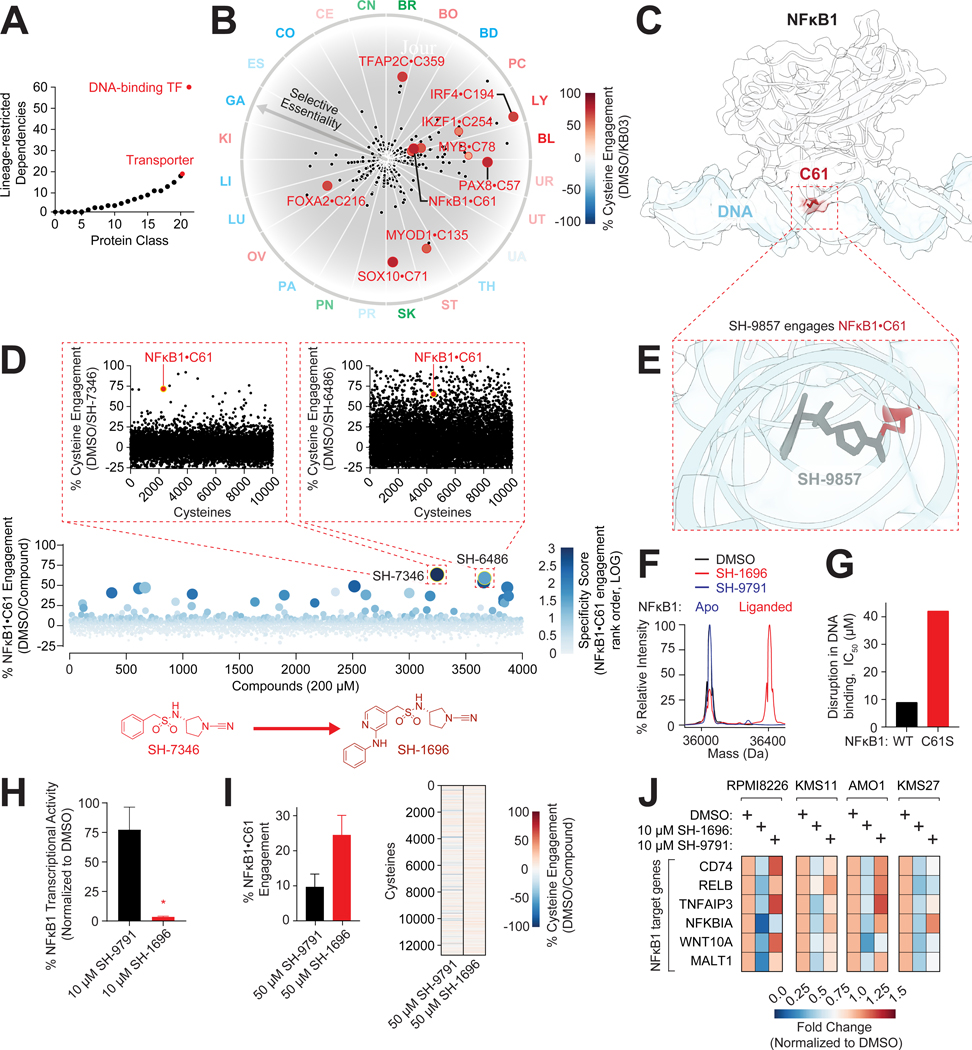
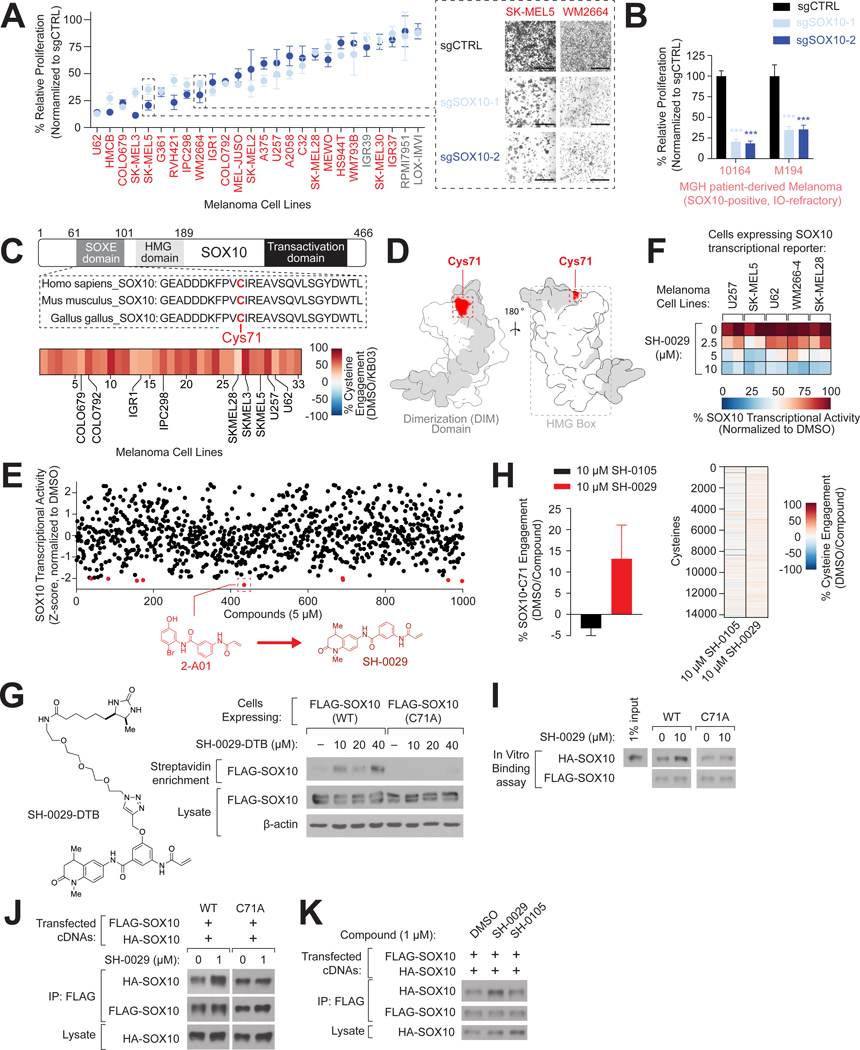
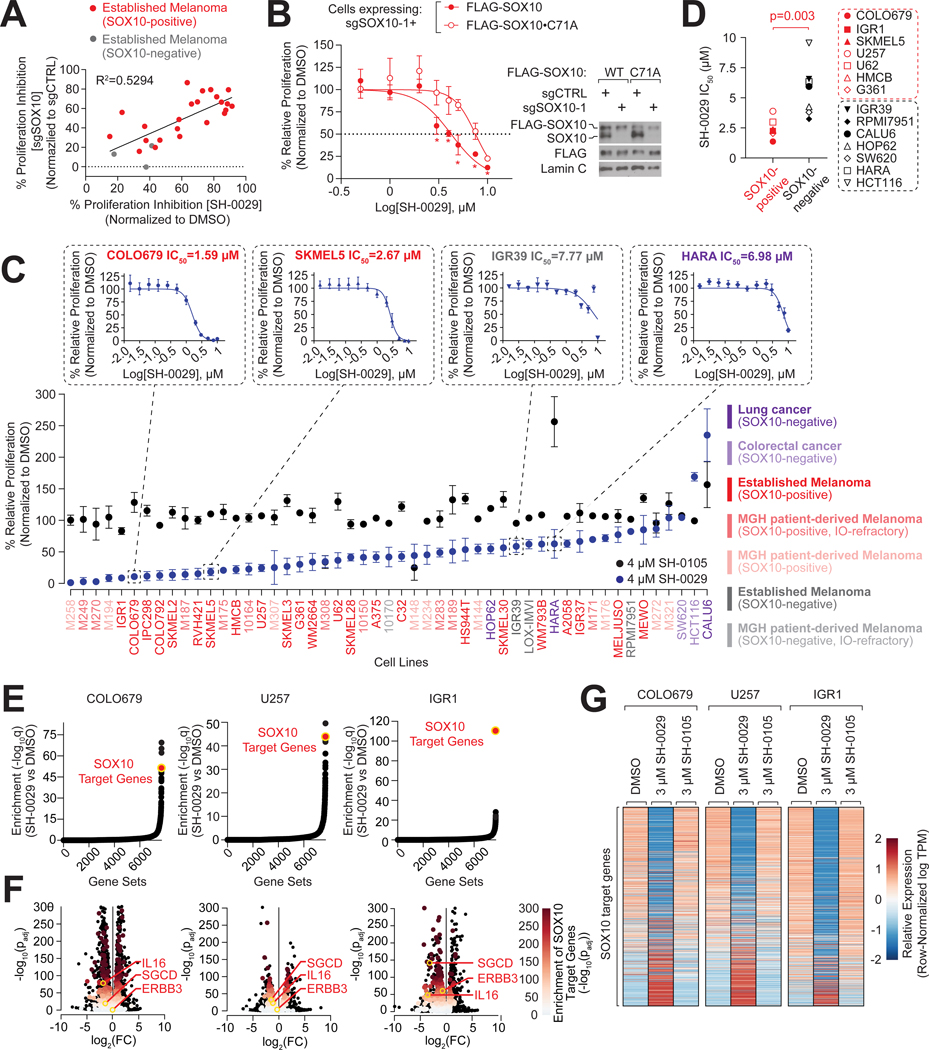
Cysteine-focused chemical proteomic platforms have accelerated the clinical development of covalent inhibitors for a wide range of targets in cancer. However, how different oncogenic contexts influence cysteine targeting remains unknown. To address this question, we have developed "DrugMap," an atlas of cysteine ligandability compiled across 416 cancer cell lines. We unexpectedly find that cysteine ligandability varies across cancer cell lines, and we attribute this to differences in cellular redox states, protein conformational changes, and genetic mutations. Leveraging these findings, we identify actionable cysteines in NF-κB1 and SOX10 and develop corresponding covalent ligands that block the activity of these transcription factors. We demonstrate that the NF-κB1 probe blocks DNA binding, whereas the SOX10 ligand increases SOX10-SOX10 interactions and disrupts melanoma transcriptional signaling. Our findings reveal heterogeneity in cysteine ligandability across cancers, pinpoint cell-intrinsic features driving cysteine targeting, and illustrate the use of covalent probes to disrupt oncogenic transcription-factor activity.
Keywords: chemical proteomics; covalent inhibitors; cysteine ligandability; transcription factors.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests D.E.F. has a financial interest in Soltego. S.M. and D.A.H. are cofounders of TellBio. G.M.B. has sponsored research agreements with Olink Proteomics, Teiko Bio, InterVenn Biosciences, and Palleon Pharmaceuticals; served on advisory boards for Iovance, Merck, Nektar Therapeutics, Novartis, and Ankyra Therapeutics; consults for Merck, InterVenn Biosciences, Iovance, and Ankyra Therapeutics; and holds equity in Ankyra Therapeutics. M.S.-F. received funding from Calico Life Sciences, Bristol-Myers Squibb, Istari Oncology, and consultants for Galvanize Therapeutics. R.W.J. is a member of the advisory board for/has a financial interest in Xsphera Biosciences Inc. R.W.J. has received honoraria from Incyte (invited speaker), G1 Therapeutics (advisory board), and Bioxcel Therapeutics (invited speaker). R.W.J. has ownership interest in U.S. patents US20200399573A9 and US20210363595A1. A.N.H. has received grants/research support from Amgen, Blueprint Medicines, BridgeBio, Bristol-Myers Squibb, C4 Therapeutics, Eli Lilly, Novartis, Nuvalent, Pfizer, Roche/Genentech, and Scorpion Therapeutics. A.N.H. has served as a compensated consultant for Amgen, Engine Biosciences, Nuvalent, Oncovalent, Pfizer, TigaTx, and Tolremo Therapeutics. M.L.S. is an equity holder and scientific co-founder/advisory board member of Immunitas Therapeutics. C.J.O. received funding from Scorpion Therapeutics, Gilead Sciences, and eFFECTOR Therapeutics. L.B.-P. is a founder/consultant/holds privately held equity in Scorpion Therapeutics. All relationships for investigators have been reviewed/managed by their respective institutions in accordance with their conflict-of-interest policies. M.M., B.H., R.D.W., H.E.M., C.-C.T., G.K., N.B., J.R.N., and B.R.M. are employees of Scorpion Therapeutics and some hold equity.
Figures
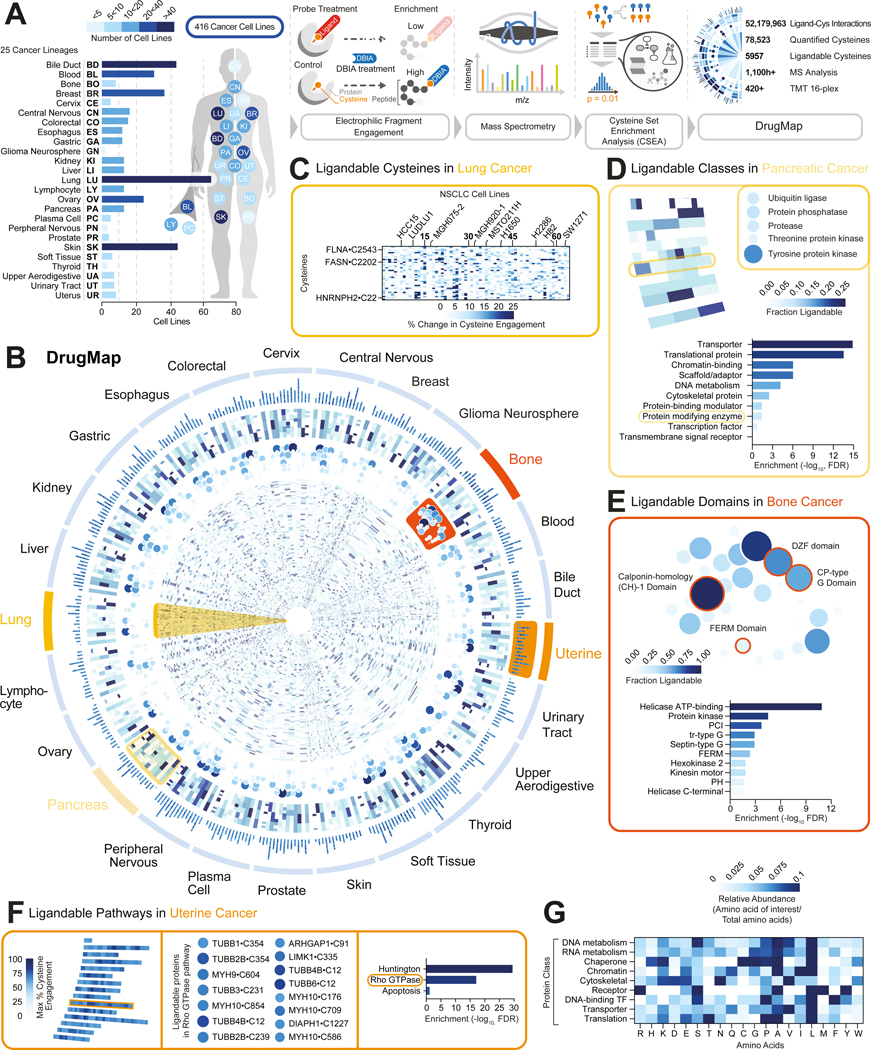
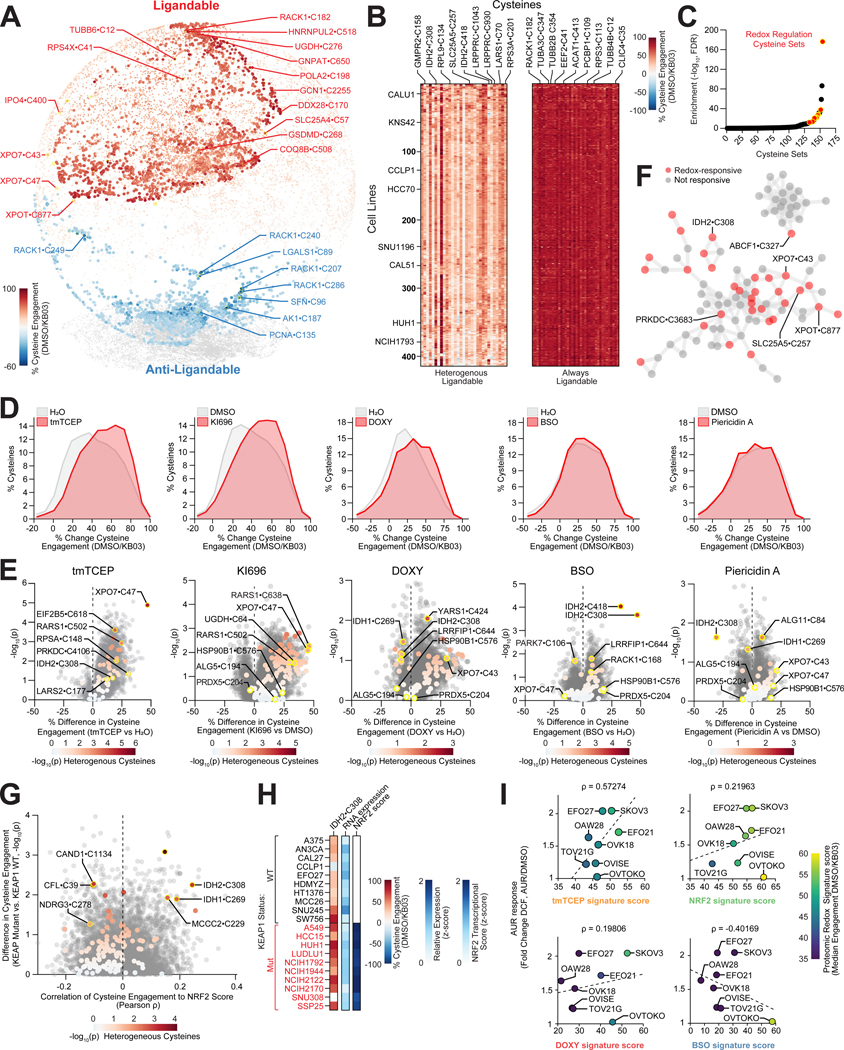
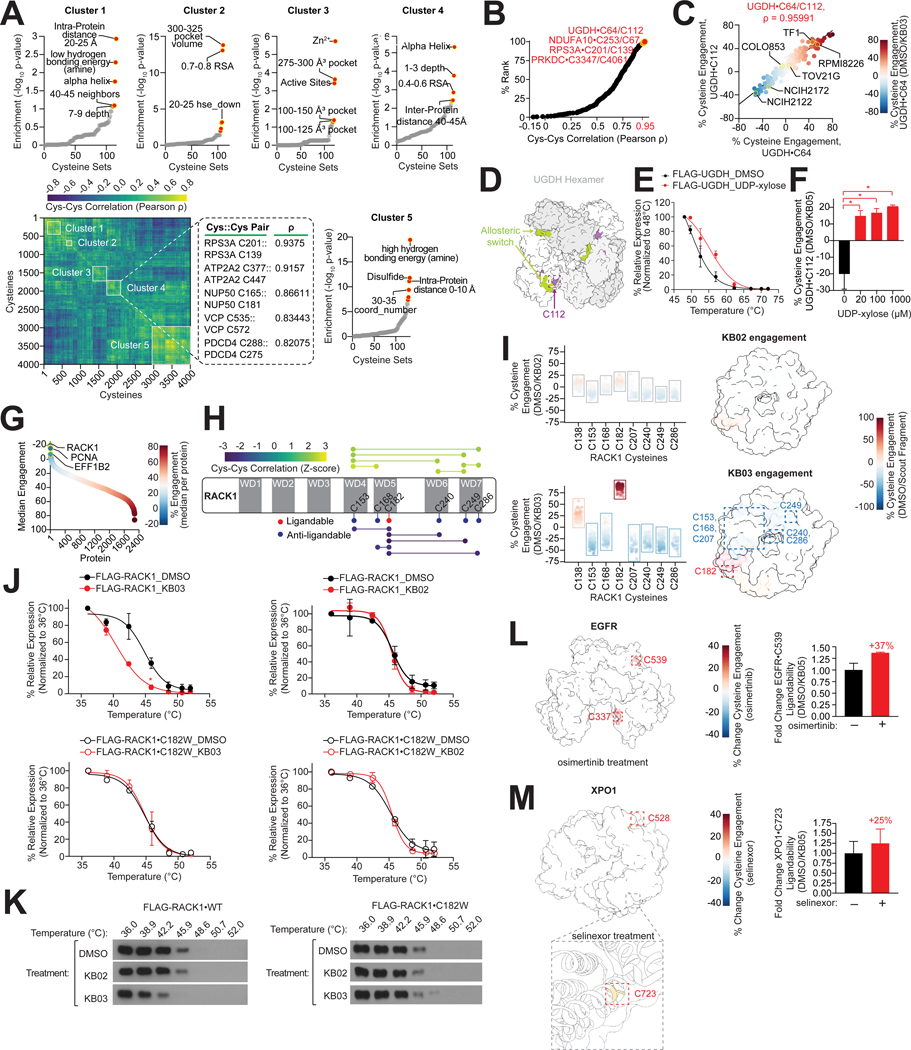
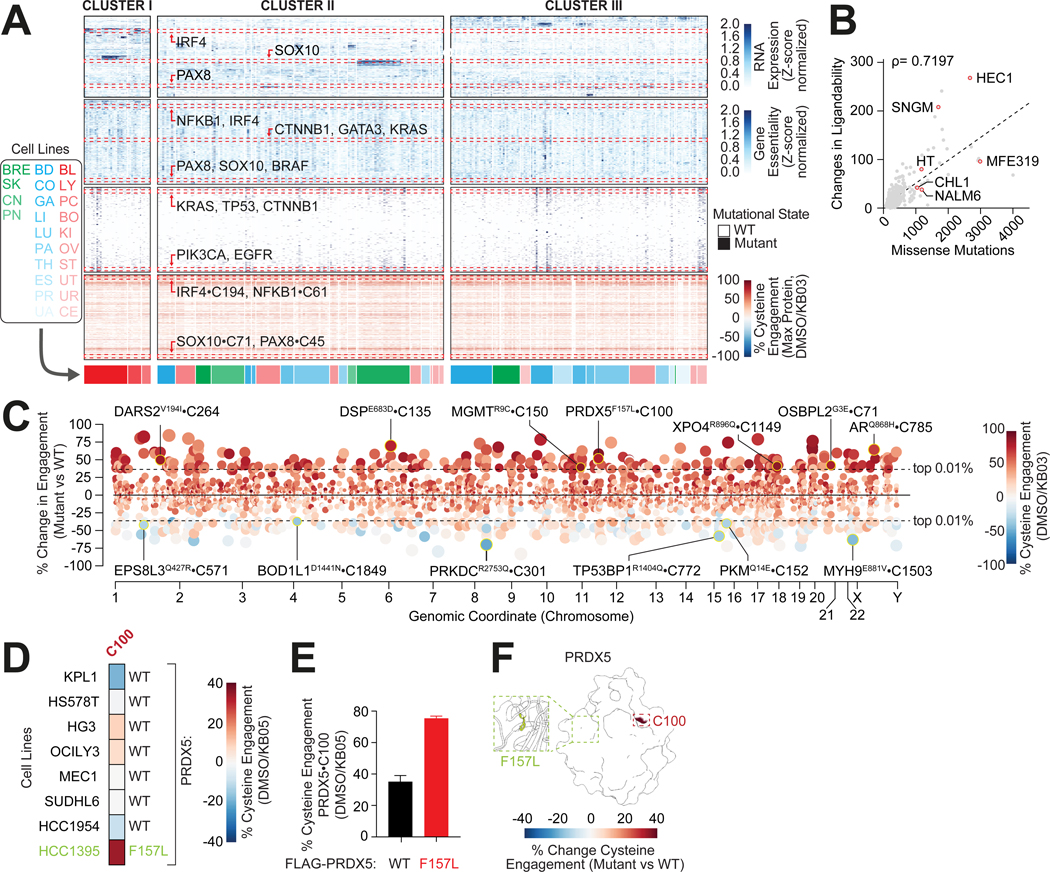
Update of
-
DrugMap: A quantitative pan-cancer analysis of cysteine ligandability.bioRxiv [Preprint]. 2023 Oct 23:2023.10.20.563287. doi: 10.1101/2023.10.20.563287. bioRxiv. 2023. Update in: Cell. 2024 May 9;187(10):2536-2556.e30. doi: 10.1016/j.cell.2024.03.027. PMID: 37961514 Free PMC article. Updated. Preprint.
Comment in
-
Mapping cysteine ligandability.Nat Rev Drug Discov. 2024 Jun;23(6):420. doi: 10.1038/d41573-024-00074-8. Nat Rev Drug Discov. 2024. PMID: 38710826 No abstract available.
References
-
- Hanahan D. (2022). Hallmarks of Cancer: New Dimensions. Cancer Discov 12, 31–46. 10.1158/2159-8290.Cd-21-1059. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- K08 CA226391/CA/NCI NIH HHS/United States
- R37 CA245523/CA/NCI NIH HHS/United States
- R21 AG083413/AG/NIA NIH HHS/United States
- K99 CA215249/CA/NCI NIH HHS/United States
- P01 CA163222/CA/NCI NIH HHS/United States
- R37 CA260062/CA/NCI NIH HHS/United States
- R21 CA256082/CA/NCI NIH HHS/United States
- DP2 GM137494/GM/NIGMS NIH HHS/United States
- R35 GM146933/GM/NIGMS NIH HHS/United States
- R35 GM153476/GM/NIGMS NIH HHS/United States
- R00 CA215249/CA/NCI NIH HHS/United States
- R01 CA137008/CA/NCI NIH HHS/United States
- R01 AR072304/AR/NIAMS NIH HHS/United States
- R01 CA129933/CA/NCI NIH HHS/United States
- R01 CA164273/CA/NCI NIH HHS/United States
- R01 CA222871/CA/NCI NIH HHS/United States
- R01 AR043369/AR/NIAMS NIH HHS/United States
- R01 CA285415/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
