

Integrated analysis of transcriptome and miRNAome reveals the heat stress response of Pinellia ternata seedlings
- PMID: 38654150
- PMCID: PMC11040748
- DOI: 10.1186/s12864-024-10318-x
Integrated analysis of transcriptome and miRNAome reveals the heat stress response of Pinellia ternata seedlings
Abstract
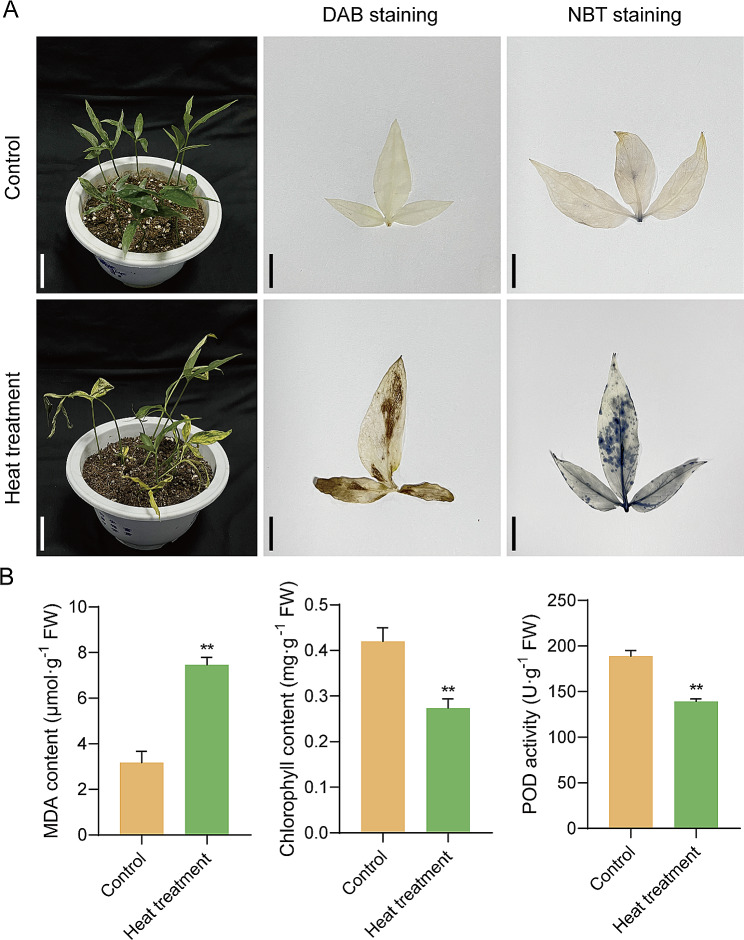
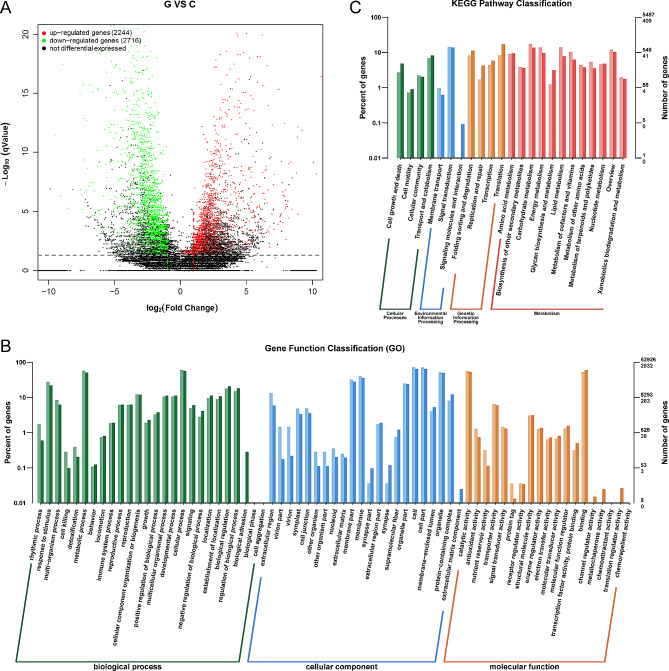
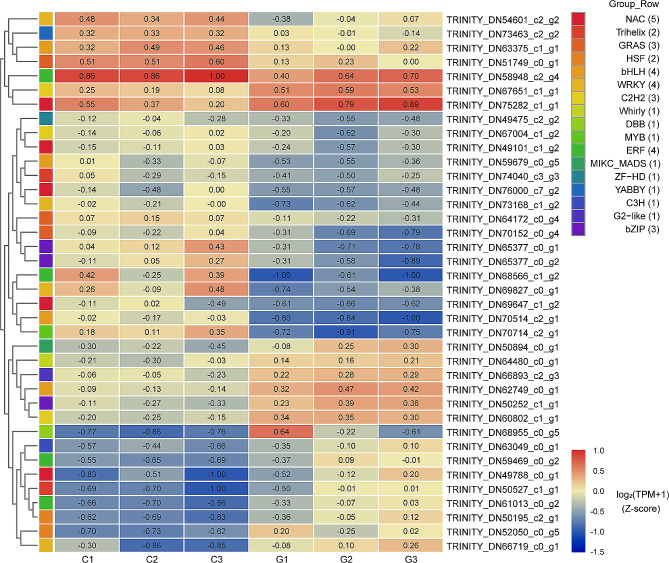
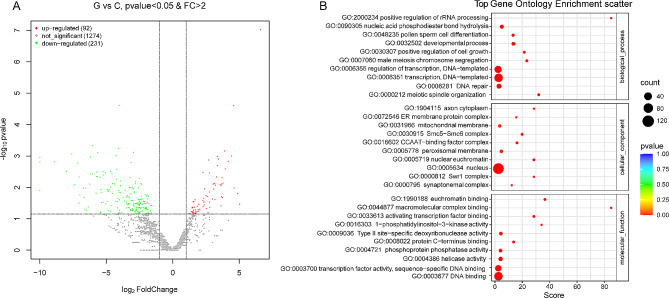
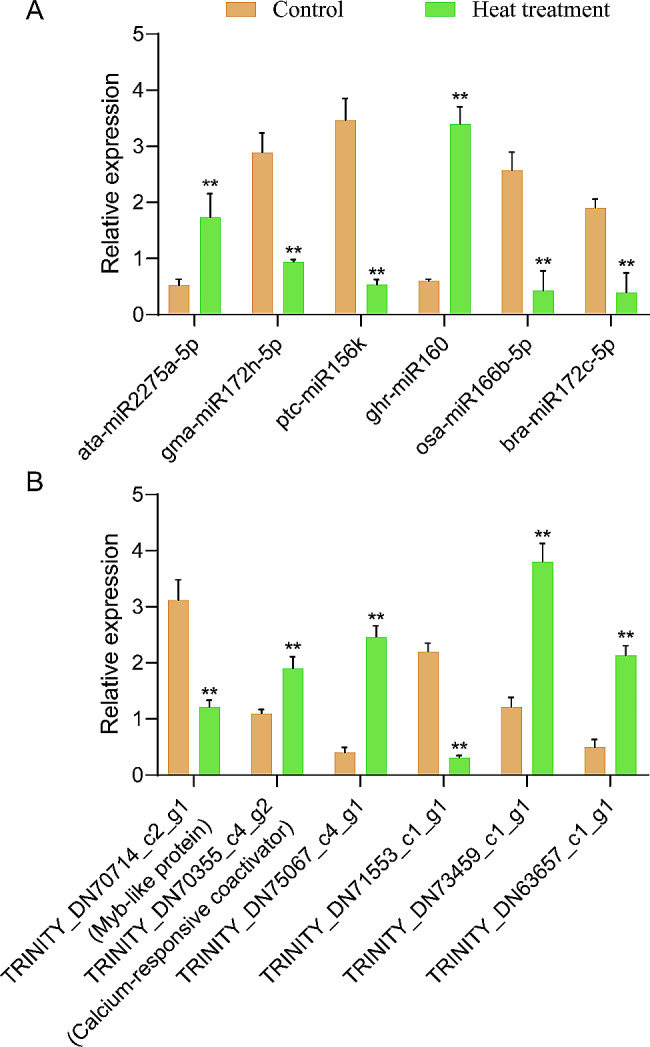
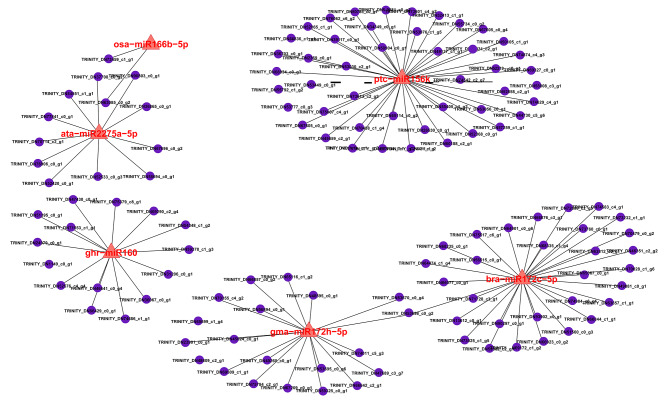
Pinellia ternata (Thunb.) Briet., a valuable herb native to China, is susceptible to the "sprout tumble" phenomenon because of high temperatures, resulting in a significant yield reduction. However, the molecular regulatory mechanisms underlying the response of P. ternata to heat stress are not well understood. In this study, we integrated transcriptome and miRNAome sequencing to identify heat-response genes, microRNAs (miRNAs), and key miRNA-target pairs in P. ternata that differed between heat-stress and room-temperature conditions. Transcriptome analysis revealed extensive reprogramming of 4,960 genes across various categories, predominantly associated with cellular and metabolic processes, responses to stimuli, biological regulation, cell parts, organelles, membranes, and catalytic and binding activities. miRNAome sequencing identified 1,597 known/conserved miRNAs that were differentially expressed between the two test conditions. According to the analysis, genes and miRNAs associated with the regulation of transcription, DNA template, transcription factor activity, and sequence-specific DNA binding pathways may play a major role in the resistance to heat stress in P. ternata. Integrated analysis of the transcriptome and miRNAome expression data revealed 41 high-confidence miRNA-mRNA pairs, forming 25 modules. MYB-like proteins and calcium-responsive transcription coactivators may play an integral role in heat-stress resistance in P. ternata. Additionally, the candidate genes and miRNAs were subjected to quantitative real-time polymerase chain reaction to validate their expression patterns. These results offer a foundation for future studies exploring the mechanisms and critical genes involved in heat-stress resistance in P. ternata.
Keywords: Pinellia ternata; Heat stress; Regulation; Transcriptome; miRNAs.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Integrated analysis of metabolome and transcriptome reveals key candidate genes involved in flavonoid biosynthesis in Pinellia ternata under heat stress.J Plant Res. 2023 May;136(3):359-369. doi: 10.1007/s10265-023-01446-8. Epub 2023 Mar 7. J Plant Res. 2023. PMID: 36881276 Free PMC article.
-
Comprehensive in silico characterization of NAC transcription factor family of Pinellia ternata and functional analysis of PtNAC66 under high-temperature tolerance in transgenic Arabidopsis thaliana.Plant Physiol Biochem. 2024 Mar;208:108539. doi: 10.1016/j.plaphy.2024.108539. Epub 2024 Mar 19. Plant Physiol Biochem. 2024. PMID: 38513515
-
Transcriptomic analysis of short-term heat stress response in Pinellia ternata provided novel insights into the improved thermotolerance by spermidine and melatonin.Ecotoxicol Environ Saf. 2020 Oct 1;202:110877. doi: 10.1016/j.ecoenv.2020.110877. Epub 2020 Jun 20. Ecotoxicol Environ Saf. 2020. PMID: 32574862
-
Pinellia ternata (Thunb.) Breit: A review of its germplasm resources, genetic diversity and active components.J Ethnopharmacol. 2020 Dec 5;263:113252. doi: 10.1016/j.jep.2020.113252. Epub 2020 Aug 13. J Ethnopharmacol. 2020. PMID: 32798614 Review.
-
A review of the research progress on Pinellia ternata (Thunb.) Breit.: Botany, traditional uses, phytochemistry, pharmacology, toxicity and quality control.Heliyon. 2023 Nov 14;9(11):e22153. doi: 10.1016/j.heliyon.2023.e22153. eCollection 2023 Nov. Heliyon. 2023. PMID: 38058630 Free PMC article. Review.
Cited by
-
Maize leaves salt-responsive genes revealed by comparative transcriptome of salt-tolerant and salt-sensitive cultivars during the seedling stage.PeerJ. 2025 Apr 10;13:e19268. doi: 10.7717/peerj.19268. eCollection 2025. PeerJ. 2025. PMID: 40226543 Free PMC article.
References
-
- Wahid A, Gelani S, Ashraf M, Foolad MR. Heat tolerance in plants: an overview. Environ Exp Bot. 2007;61(3):199–223. doi: 10.1016/j.envexpbot.2007.05.011. - DOI
MeSH terms
Substances
Grants and funding
- 2023AH050352/Project of Natural Science Research of Universities in Anhui Province, China
- 82373993/National Natural Science Foundation of China
- 82274048/National Natural Science Foundation of China
- 2019GNC106113/Key Research and Development Program of Shandong Province, China
- 2022AH010029/Excellent Scientific Research and Innovation Team of University in Anhui Province
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
