

Diabetes Primes Neutrophils for Neutrophil Extracellular Trap Formation through Trained Immunity
- PMID: 38654733
- PMCID: PMC11037460
- DOI: 10.34133/research.0365
Diabetes Primes Neutrophils for Neutrophil Extracellular Trap Formation through Trained Immunity
Abstract
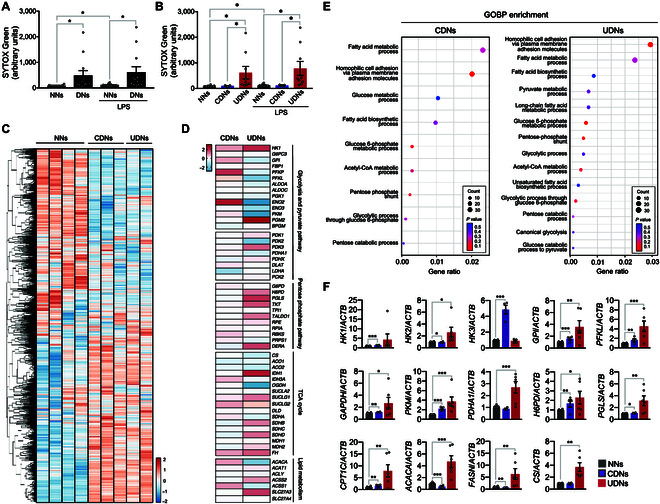
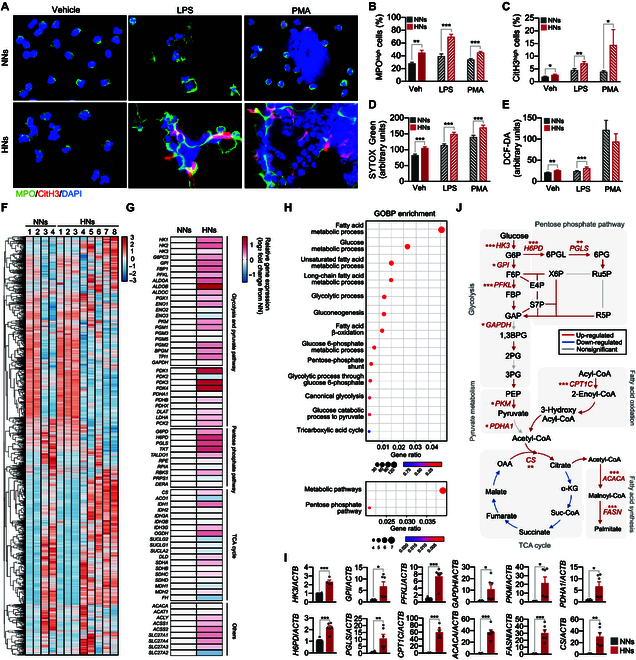
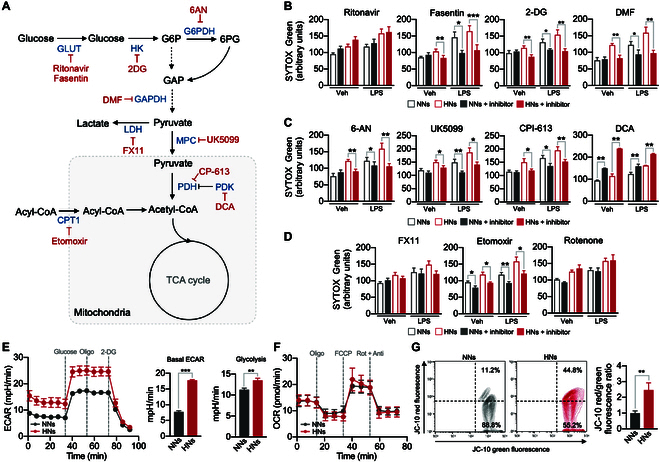
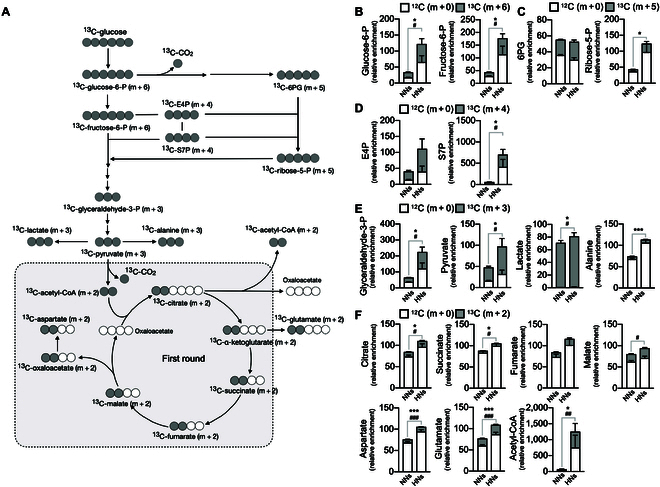
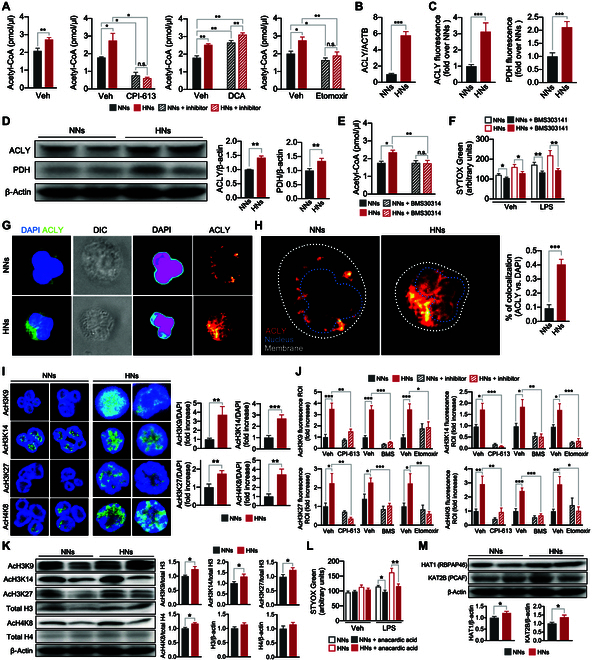
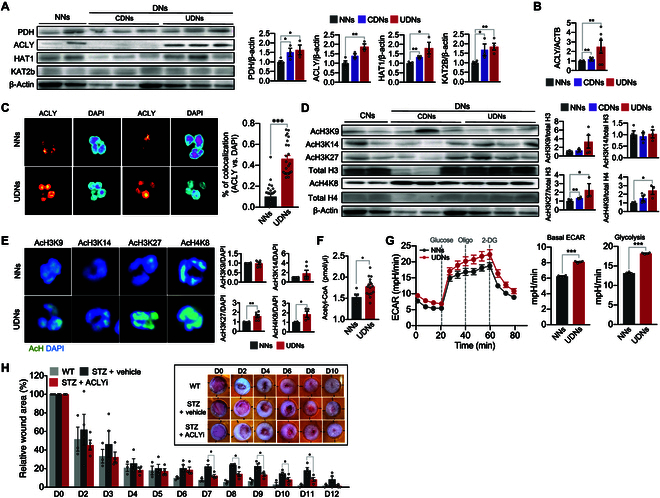
Neutrophils are primed for neutrophil extracellular trap (NET) formation during diabetes, and excessive NET formation from primed neutrophils compromises wound healing in patients with diabetes. Here, we demonstrate that trained immunity mediates diabetes-induced NET priming in neutrophils. Under diabetic conditions, neutrophils exhibit robust metabolic reprogramming comprising enhanced glycolysis via the pentose phosphate pathway and fatty acid oxidation, which result in the accumulation of acetyl-coenzyme A. Adenosine 5'-triphosphate-citrate lyase-mediated accumulation of acetyl-coenzyme A and histone acetyltransferases further induce the acetylation of lysine residues on histone 3 (AcH3K9, AcH3K14, and AcH3K27) and histone 4 (AcH4K8). The pharmacological inhibition of adenosine 5'-triphosphate-citrate lyase and histone acetyltransferases completely inhibited high-glucose-induced NET priming. The trained immunity of neutrophils was further confirmed in neutrophils isolated from patients with diabetes. Our findings suggest that trained immunity mediates functional changes in neutrophils in diabetic environments, and targeting neutrophil-trained immunity may be a potential therapeutic target for controlling inflammatory complications of diabetes.
Copyright © 2024 Sanjeeb Shrestha et al.
Conflict of interest statement
Competing interests: The authors declare that they have no competing interests.
Figures
References
LinkOut - more resources
Full Text Sources
