

Goal-directed and flexible modulation of syllable sequence within birdsong
- PMID: 38658545
- PMCID: PMC11043396
- DOI: 10.1038/s41467-024-47824-1
Goal-directed and flexible modulation of syllable sequence within birdsong
Abstract
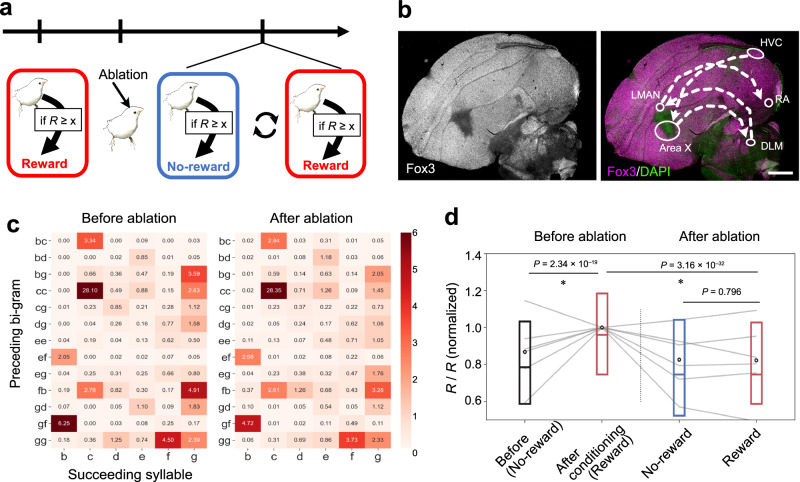
Songs constitute a complex system of vocal signals for inter-individual communication in songbirds. Here, we elucidate the flexibility which songbirds exhibit in the organizing and sequencing of syllables within their songs. Utilizing a newly devised song decoder for quasi-real-time annotation, we execute an operant conditioning paradigm, with rewards contingent upon specific syllable syntax. Our analysis reveals that birds possess the capacity to modify the contents of their songs, adjust the repetition length of particular syllables and employing specific motifs. Notably, birds altered their syllable sequence in a goal-directed manner to obtain rewards. We demonstrate that such modulation occurs within a distinct song segment, with adjustments made within 10 minutes after cue presentation. Additionally, we identify the involvement of the parietal-basal ganglia pathway in orchestrating these flexible modulations of syllable sequences. Our findings unveil an unappreciated aspect of songbird communication, drawing parallels with human speech.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
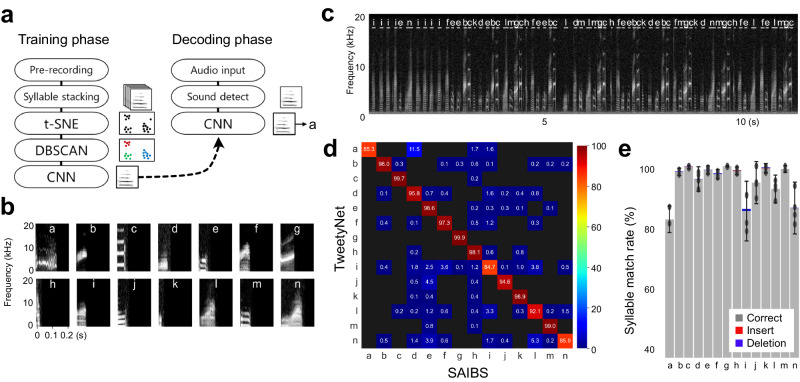
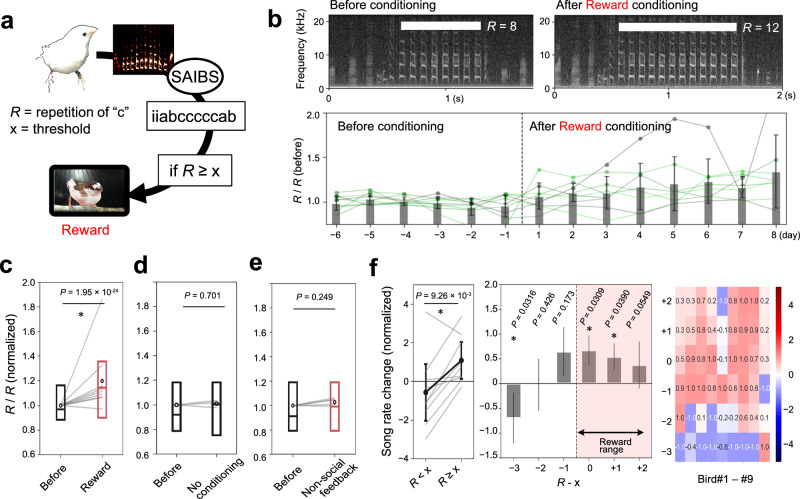
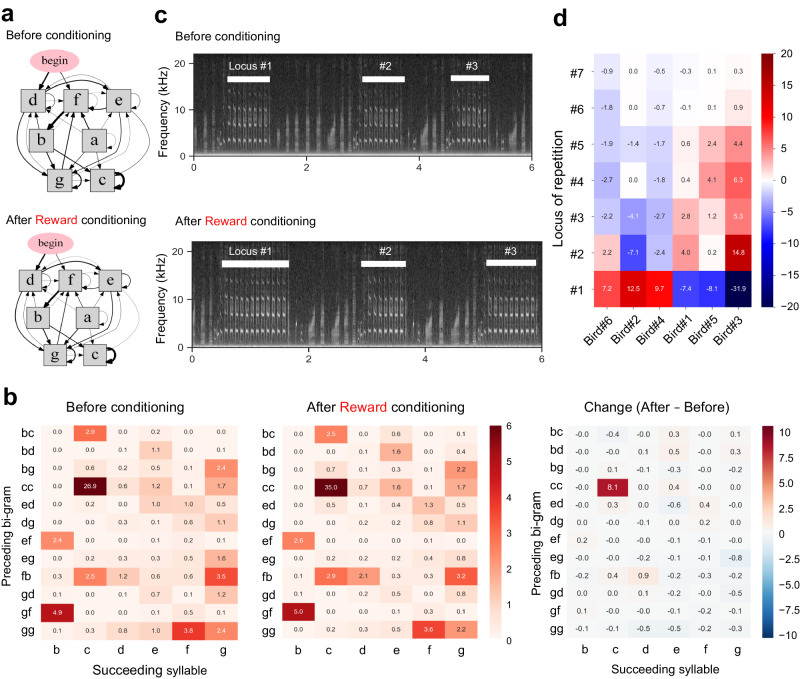
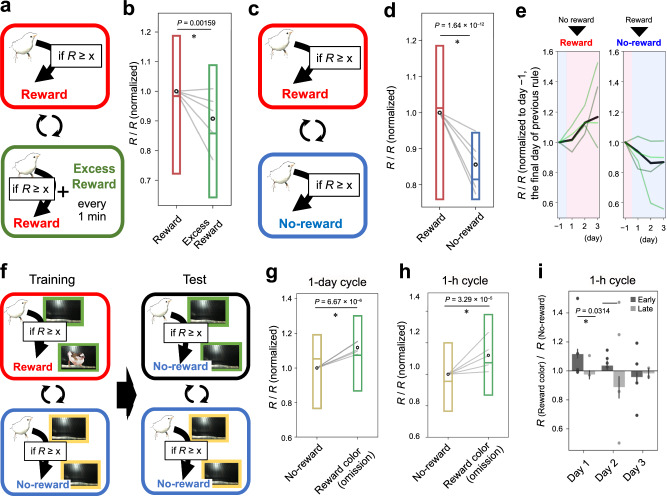

Similar articles
-
Songbirds can learn flexible contextual control over syllable sequencing.Elife. 2021 Jun 1;10:e61610. doi: 10.7554/eLife.61610. Elife. 2021. PMID: 34060473 Free PMC article.
-
Sound sequences in birdsong: how much do birds really care?Philos Trans R Soc Lond B Biol Sci. 2020 Jan 6;375(1789):20190044. doi: 10.1098/rstb.2019.0044. Epub 2019 Nov 18. Philos Trans R Soc Lond B Biol Sci. 2020. PMID: 31735149 Free PMC article. Review.
-
Do songbirds hear songs syllable by syllable?Behav Processes. 2020 May;174:104089. doi: 10.1016/j.beproc.2020.104089. Epub 2020 Feb 24. Behav Processes. 2020. PMID: 32105758
-
Social modulation of sequence and syllable variability in adult birdsong.J Neurophysiol. 2008 Apr;99(4):1700-11. doi: 10.1152/jn.01296.2007. Epub 2008 Jan 23. J Neurophysiol. 2008. PMID: 18216221
-
Birdsong and the brain: the syntax of memory.Neuroreport. 2010 Apr 21;21(6):395-8. doi: 10.1097/WNR.0b013e3283385949. Neuroreport. 2010. PMID: 20220539 Review.
Cited by
-
A behavioral analysis system MCFBM enables objective inference of songbirds' attention during social interactions.Cell Rep Methods. 2024 Sep 16;4(9):100844. doi: 10.1016/j.crmeth.2024.100844. Epub 2024 Sep 3. Cell Rep Methods. 2024. PMID: 39232558 Free PMC article.
-
Song determined by phylogeny and body mass in two differently constrained groups of birds: manakins and cardinals.BMC Ecol Evol. 2024 Aug 19;24(1):109. doi: 10.1186/s12862-024-02298-z. BMC Ecol Evol. 2024. PMID: 39160456 Free PMC article.
-
Social context affects sequence modification learning in birdsong.Front Psychol. 2025 Feb 5;16:1488762. doi: 10.3389/fpsyg.2025.1488762. eCollection 2025. Front Psychol. 2025. PMID: 39973966 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
