

Transient loss of Polycomb components induces an epigenetic cancer fate
- PMID: 38658752
- PMCID: PMC11096130
- DOI: 10.1038/s41586-024-07328-w
Transient loss of Polycomb components induces an epigenetic cancer fate
Abstract
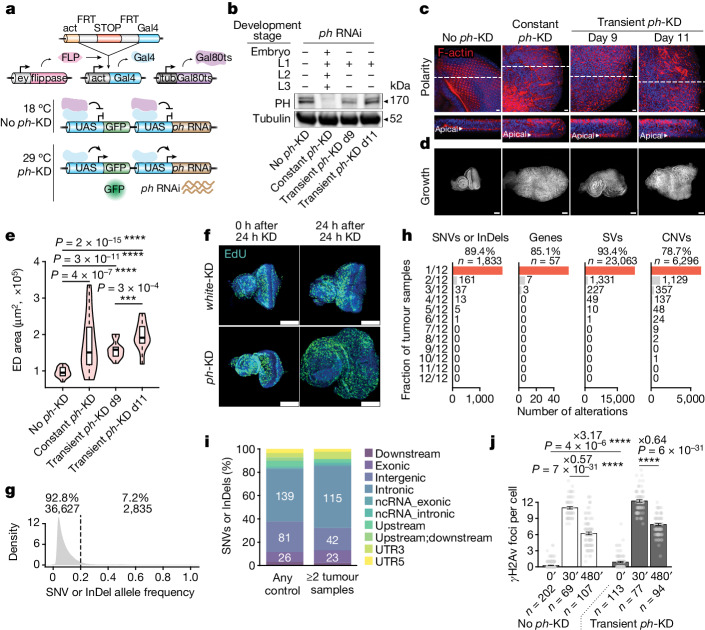
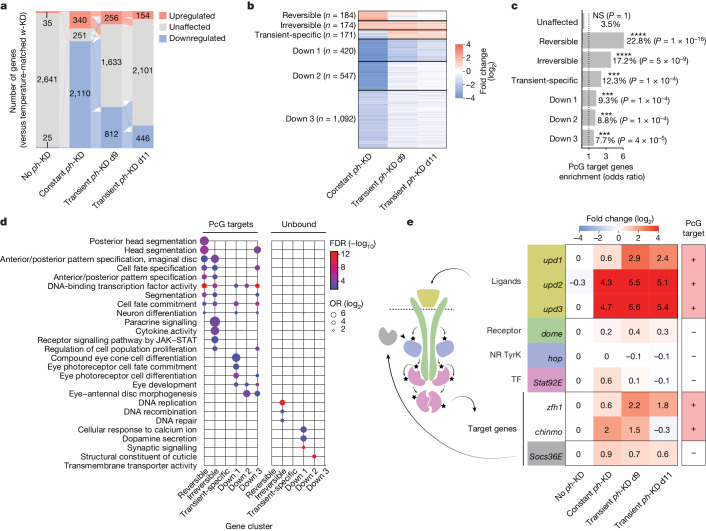
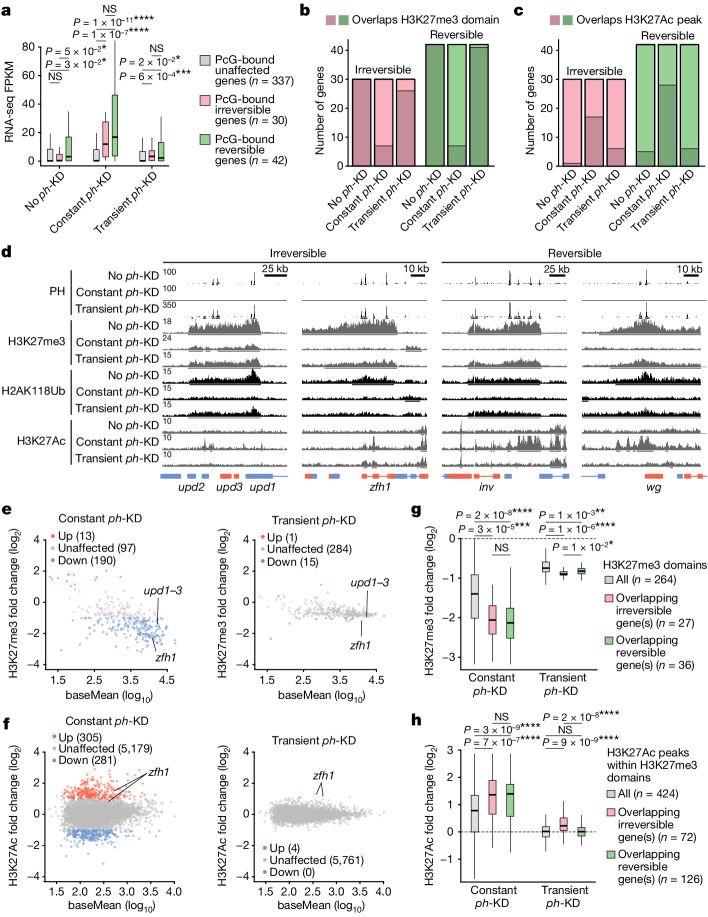
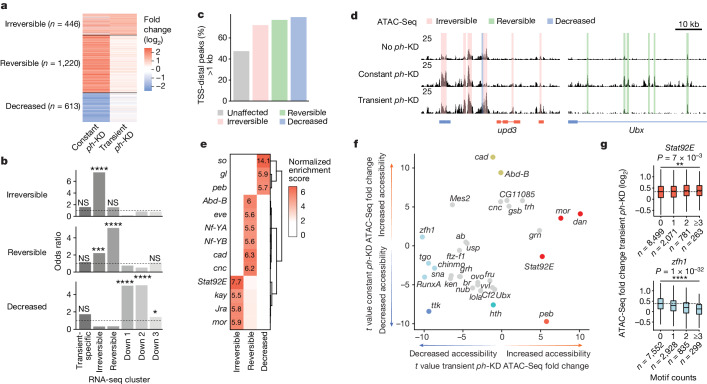
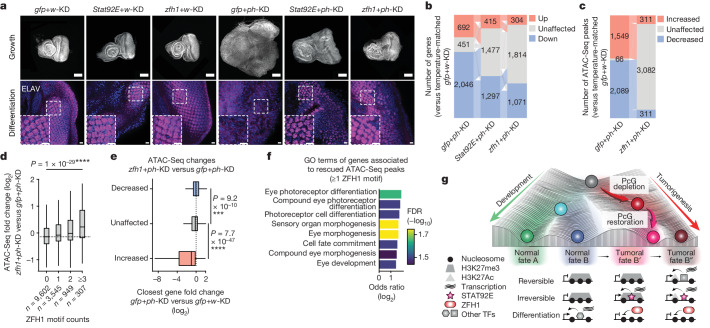
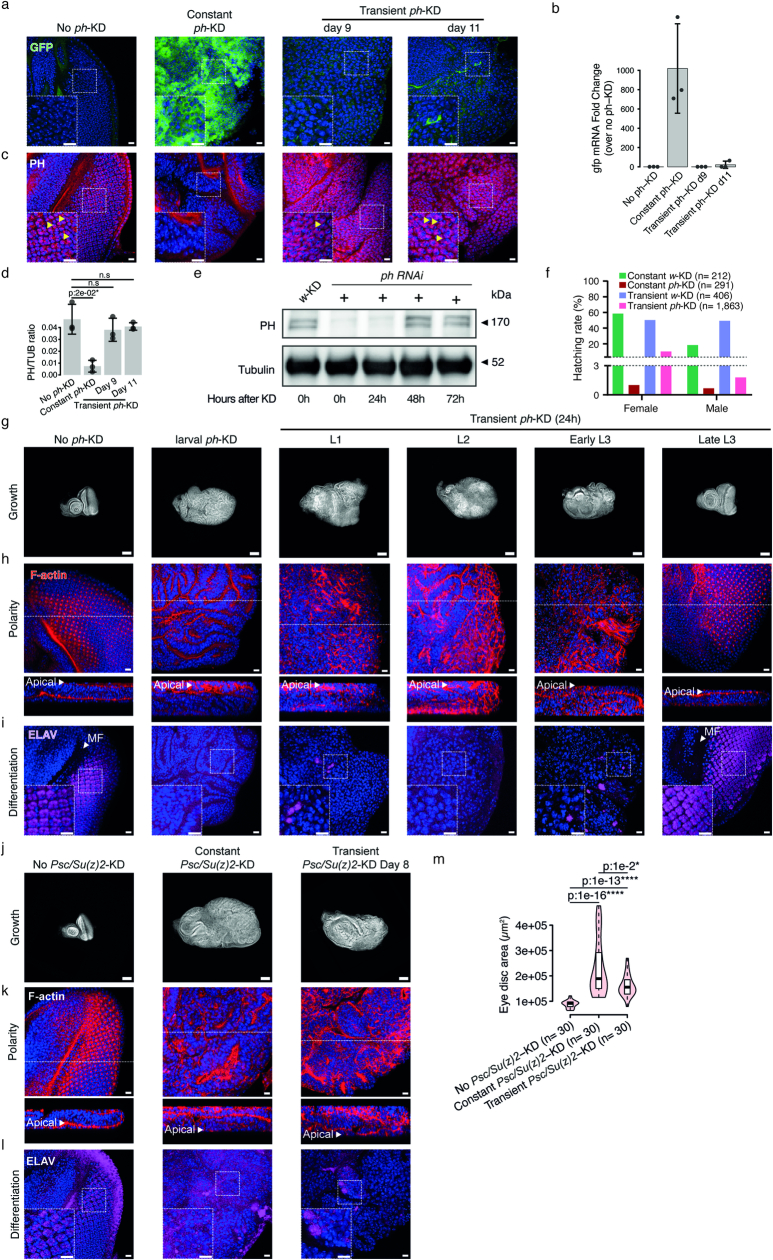
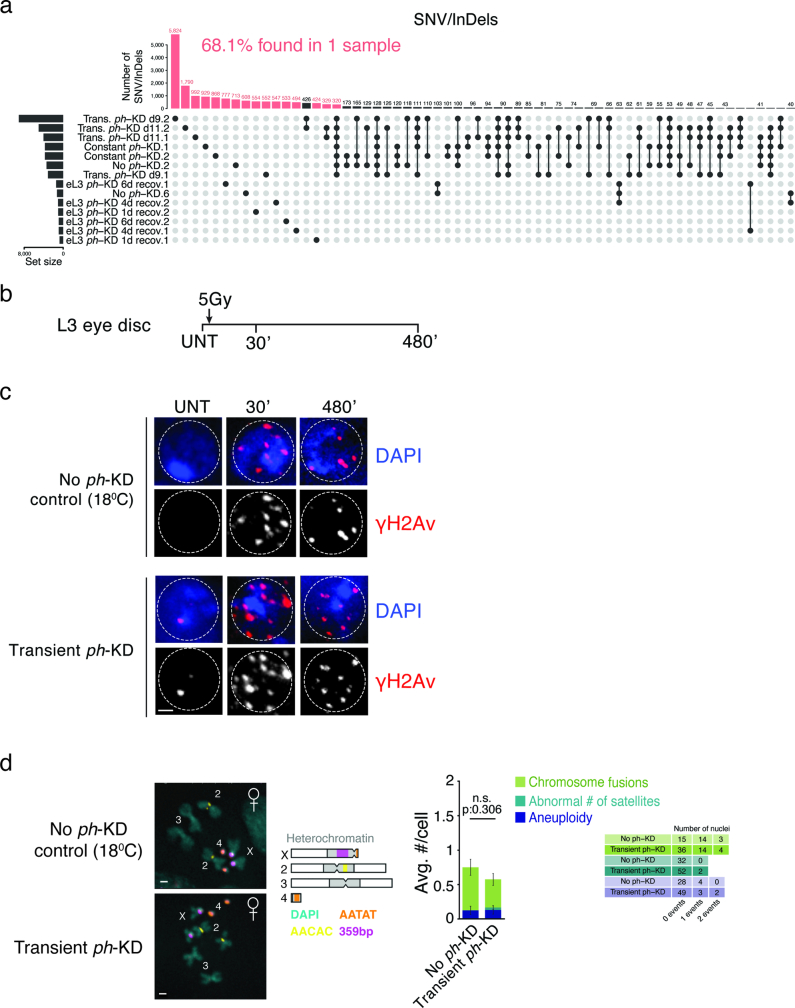
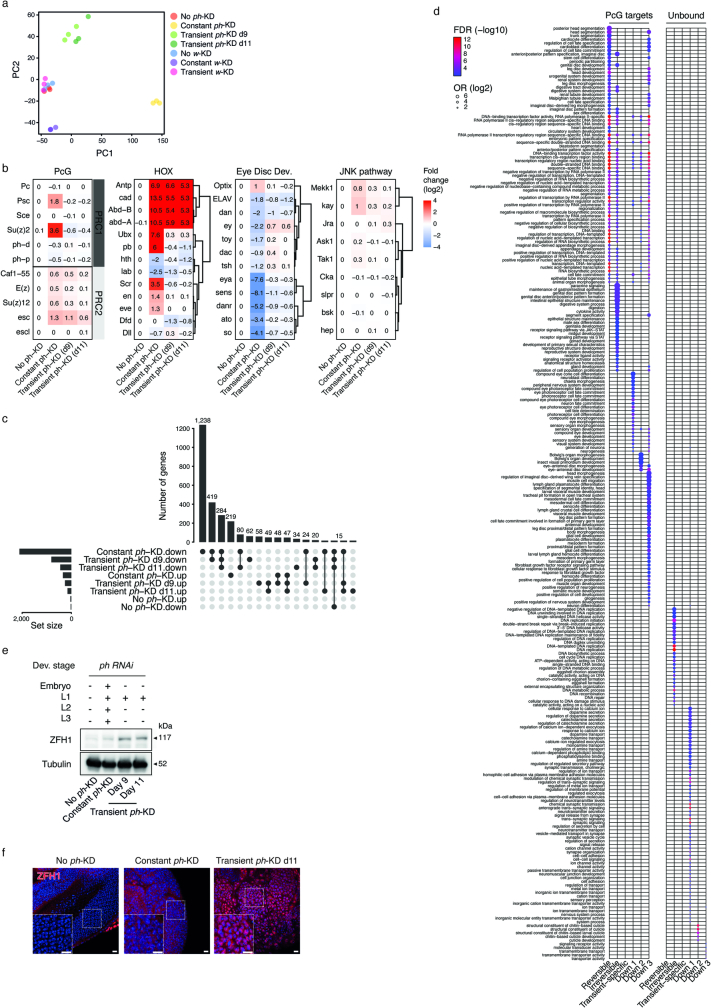
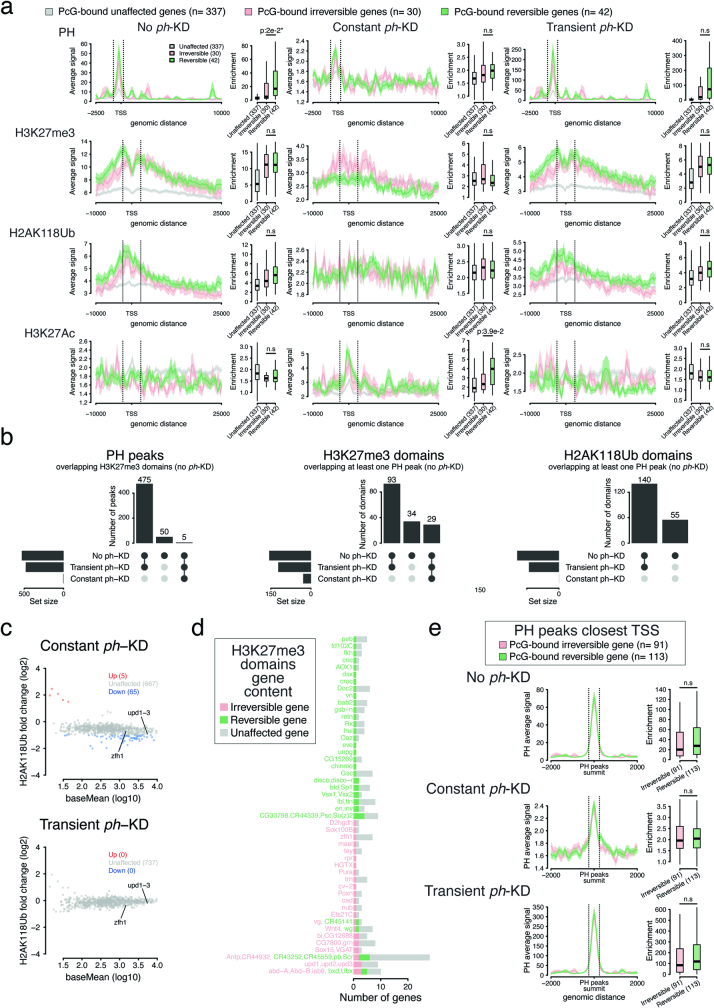
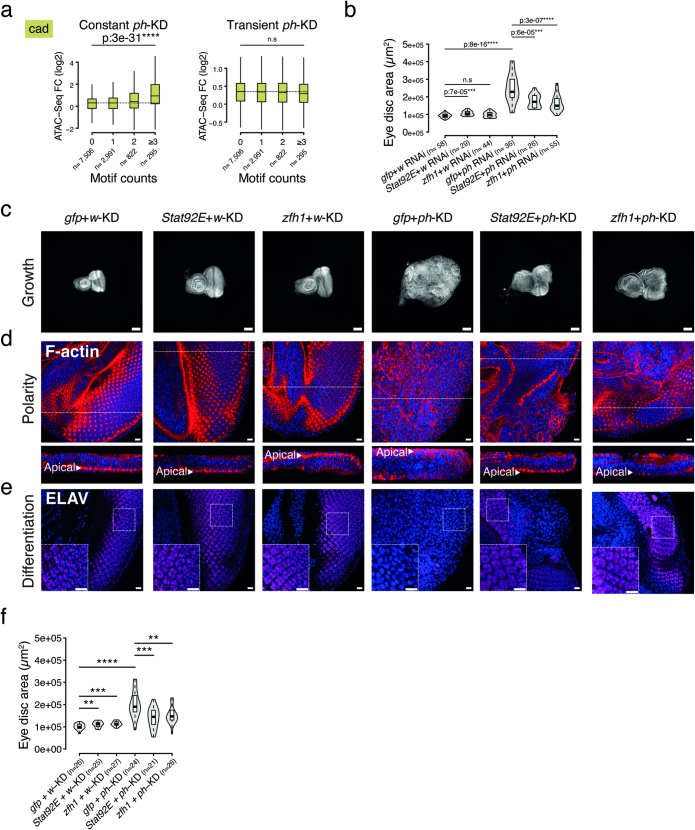
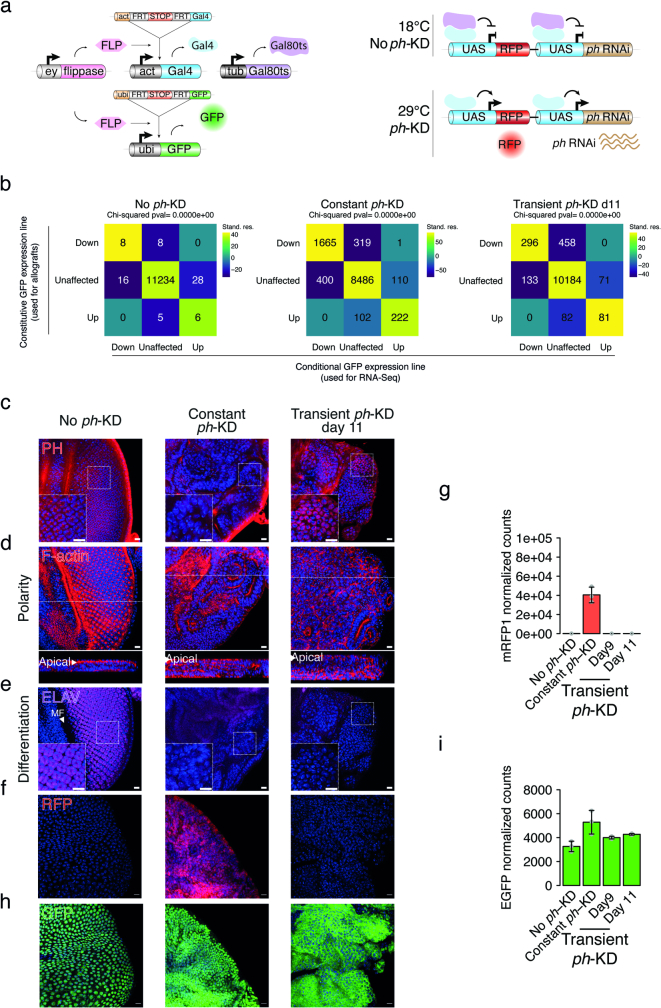
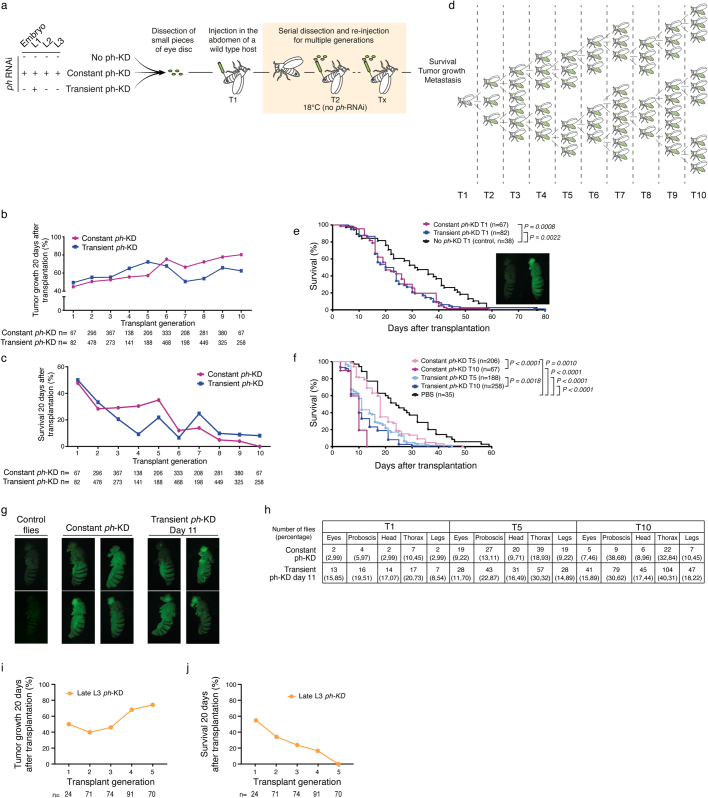
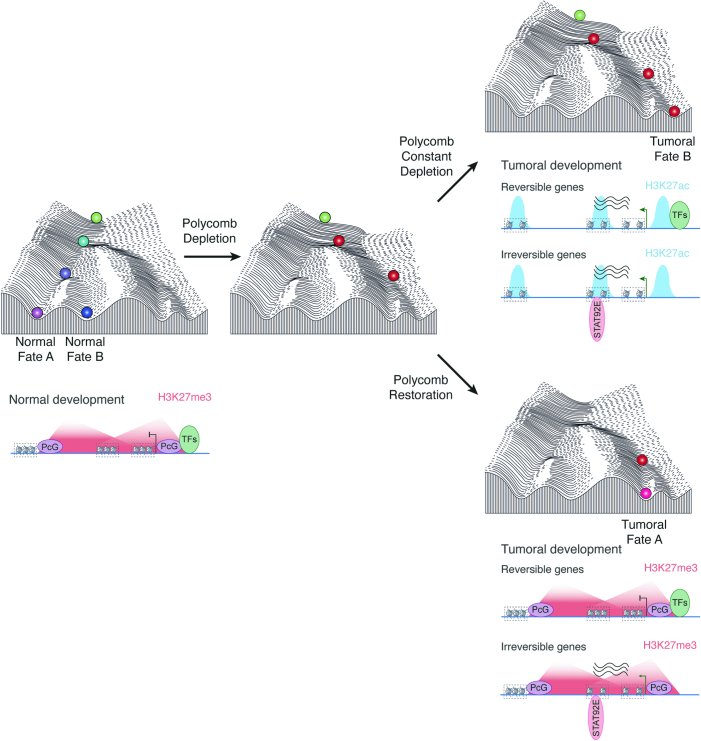
Although cancer initiation and progression are generally associated with the accumulation of somatic mutations1,2, substantial epigenomic alterations underlie many aspects of tumorigenesis and cancer susceptibility3-6, suggesting that genetic mechanisms might not be the only drivers of malignant transformation7. However, whether purely non-genetic mechanisms are sufficient to initiate tumorigenesis irrespective of mutations has been unknown. Here, we show that a transient perturbation of transcriptional silencing mediated by Polycomb group proteins is sufficient to induce an irreversible switch to a cancer cell fate in Drosophila. This is linked to the irreversible derepression of genes that can drive tumorigenesis, including members of the JAK-STAT signalling pathway and zfh1, the fly homologue of the ZEB1 oncogene, whose aberrant activation is required for Polycomb perturbation-induced tumorigenesis. These data show that a reversible depletion of Polycomb proteins can induce cancer in the absence of driver mutations, suggesting that tumours can emerge through epigenetic dysregulation leading to inheritance of altered cell fates.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
