

PLC-γ-Ca2+ pathway regulates axonal TrkB endocytosis and is required for long-distance propagation of BDNF signaling
- PMID: 38660384
- PMCID: PMC11040097
- DOI: 10.3389/fnmol.2024.1009404
PLC-γ-Ca2+ pathway regulates axonal TrkB endocytosis and is required for long-distance propagation of BDNF signaling
Abstract
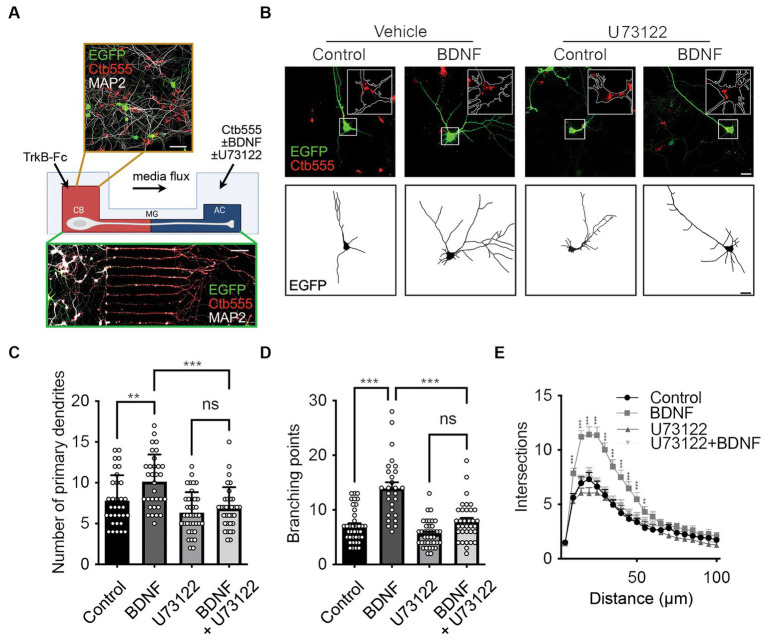
Brain-derived neurotrophic factor (BDNF) and its tropomyosin receptor kinase B (TrkB) are important signaling proteins that regulate dendritic growth and maintenance in the central nervous system (CNS). After binding of BDNF, TrkB is endocytosed into endosomes and continues signaling within the cell soma, dendrites, and axon. In previous studies, we showed that BDNF signaling initiated in axons triggers long-distance signaling, inducing dendritic arborization in a CREB-dependent manner in cell bodies, processes that depend on axonal dynein and TrkB activities. The binding of BDNF to TrkB triggers the activation of different signaling pathways, including the ERK, PLC-γ and PI3K-mTOR pathways, to induce dendritic growth and synaptic plasticity. How TrkB downstream pathways regulate long-distance signaling is unclear. Here, we studied the role of PLC-γ-Ca2+ in BDNF-induced long-distance signaling using compartmentalized microfluidic cultures. We found that dendritic branching and CREB phosphorylation induced by axonal BDNF stimulation require the activation of PLC-γ in the axons of cortical neurons. Locally, in axons, BDNF increases PLC-γ phosphorylation and induces intracellular Ca2+ waves in a PLC-γ-dependent manner. In parallel, we observed that BDNF-containing signaling endosomes transport to the cell body was dependent on PLC-γ activity and intracellular Ca2+ stores. Furthermore, the activity of PLC-γ is required for BDNF-dependent TrkB endocytosis, suggesting a role for the TrkB/PLC-γ signaling pathway in axonal signaling endosome formation.
Keywords: PLC-γ; TrkB; axonal transport; brain-derived neurotrophic factor; calcium; dendritic branching; endocytosis; signaling endosomes.
Copyright © 2024 Moya-Alvarado, Valero-Peña, Aguirre-Soto, Bustos, Lazo and Bronfman.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






Similar articles
-
BDNF/TrkB signaling endosomes in axons coordinate CREB/mTOR activation and protein synthesis in the cell body to induce dendritic growth in cortical neurons.Elife. 2023 Feb 24;12:e77455. doi: 10.7554/eLife.77455. Elife. 2023. PMID: 36826992 Free PMC article.
-
BDNF regulates Rab11-mediated recycling endosome dynamics to induce dendritic branching.J Neurosci. 2013 Apr 3;33(14):6112-22. doi: 10.1523/JNEUROSCI.4630-12.2013. J Neurosci. 2013. PMID: 23554492 Free PMC article.
-
The Rab11-regulated endocytic pathway and BDNF/TrkB signaling: Roles in plasticity changes and neurodegenerative diseases.Neurobiol Dis. 2022 Sep;171:105796. doi: 10.1016/j.nbd.2022.105796. Epub 2022 Jun 18. Neurobiol Dis. 2022. PMID: 35728773 Review.
-
c-Abl Tyrosine Kinase Is Required for BDNF-Induced Dendritic Branching and Growth.Int J Mol Sci. 2023 Jan 18;24(3):1944. doi: 10.3390/ijms24031944. Int J Mol Sci. 2023. PMID: 36768268 Free PMC article.
-
BDNF-induced local protein synthesis and synaptic plasticity.Neuropharmacology. 2014 Jan;76 Pt C:639-56. doi: 10.1016/j.neuropharm.2013.04.005. Epub 2013 Apr 16. Neuropharmacology. 2014. PMID: 23602987 Review.
Cited by
-
The Progress in Exploratory Studies of Peripheral Blood Single Nucleated Cells as Seed Cells in Peripheral Nerve Repair.Neural Plast. 2025 Aug 7;2025:8895041. doi: 10.1155/np/8895041. eCollection 2025. Neural Plast. 2025. PMID: 40822885 Free PMC article. Review.
-
Dissection of signaling pathways regulating TrkB-dependent gephyrin clustering.Front Mol Neurosci. 2024 Oct 24;17:1480820. doi: 10.3389/fnmol.2024.1480820. eCollection 2024. Front Mol Neurosci. 2024. PMID: 39534513 Free PMC article.
-
BDNF/proBDNF Interplay in the Mediation of Neuronal Apoptotic Mechanisms in Neurodegenerative Diseases.Int J Mol Sci. 2025 May 21;26(10):4926. doi: 10.3390/ijms26104926. Int J Mol Sci. 2025. PMID: 40430064 Free PMC article. Review.
References
-
- Beattie E. C., Zhou J., Grimes M. L., Bunnett N. W., Howe C. L., Mobley W. C. (1996). A signaling endosome hypothesis to explain Ngf actions: potential implications for neurodegeneration. Cold Spring Harb. Symp. Quant. Biol. 61, 389–406. PMID: - PubMed
-
- Bleasdale J. E., Thakur N. R., Gremban R. S., Bundy G. L., Fitzpatrick F. A., Smith R. J., et al. . (1990). Selective inhibition of receptor-coupled phospholipase C-dependent processes in human platelets and polymorphonuclear neutrophils. J. Pharmacol. Exp. Ther. 255, 756–768. PMID: - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous