

Dysfunction of the adhesion G protein-coupled receptor latrophilin 1 (ADGRL1/LPHN1) increases the risk of obesity
- PMID: 38664368
- PMCID: PMC11045723
- DOI: 10.1038/s41392-024-01810-7
Dysfunction of the adhesion G protein-coupled receptor latrophilin 1 (ADGRL1/LPHN1) increases the risk of obesity
Abstract
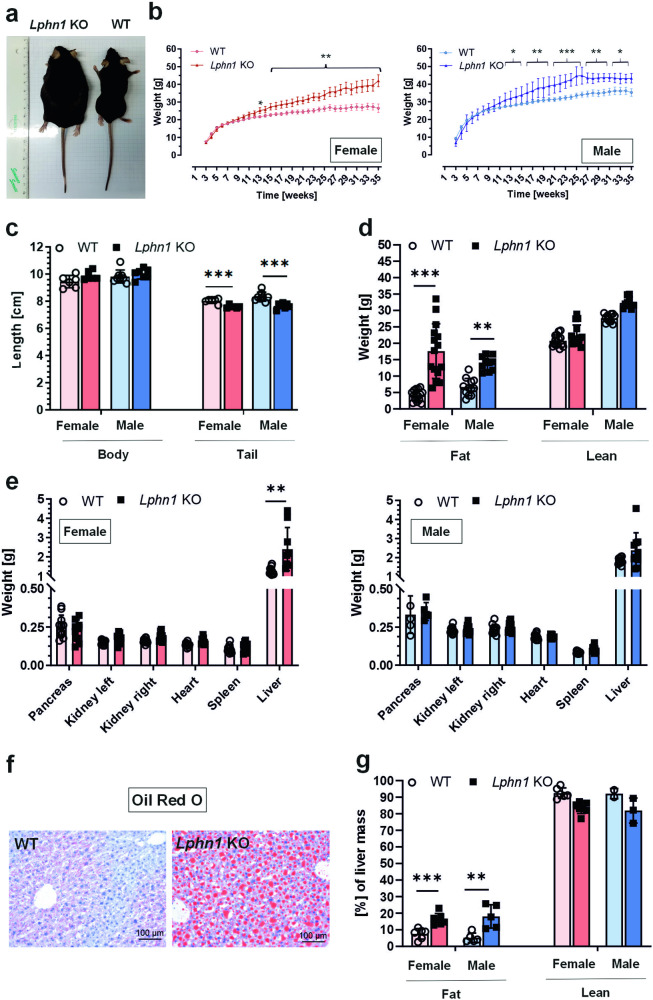
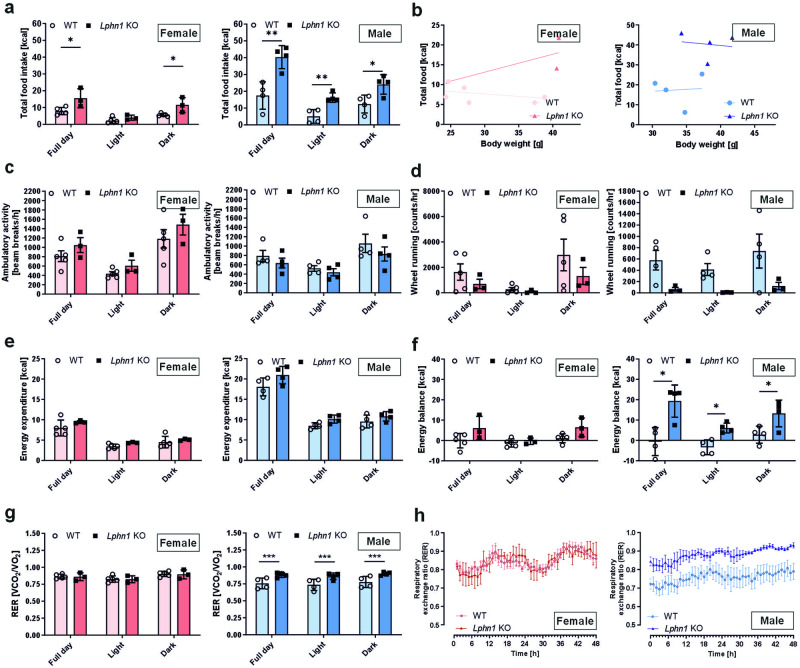
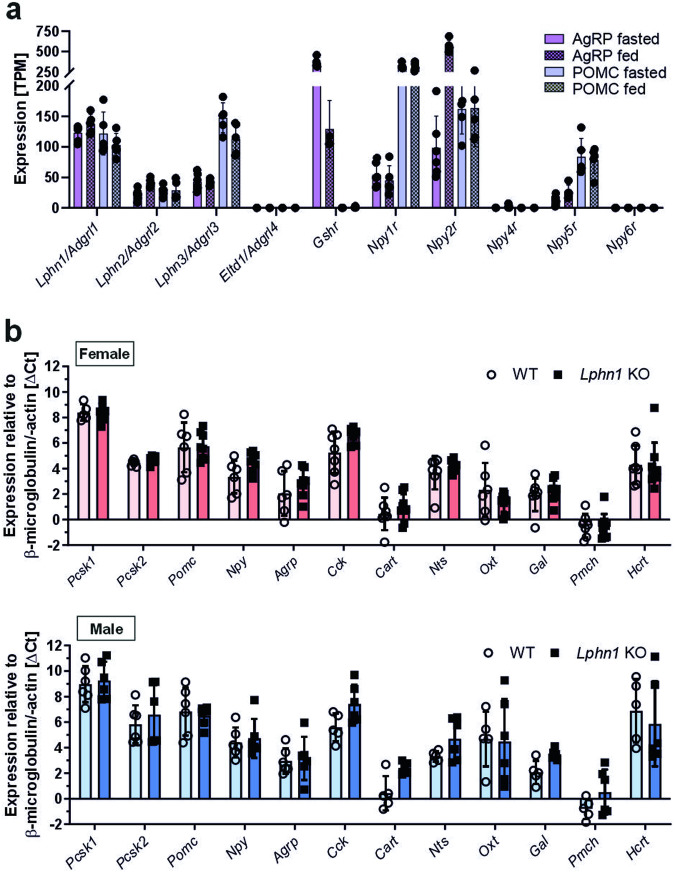
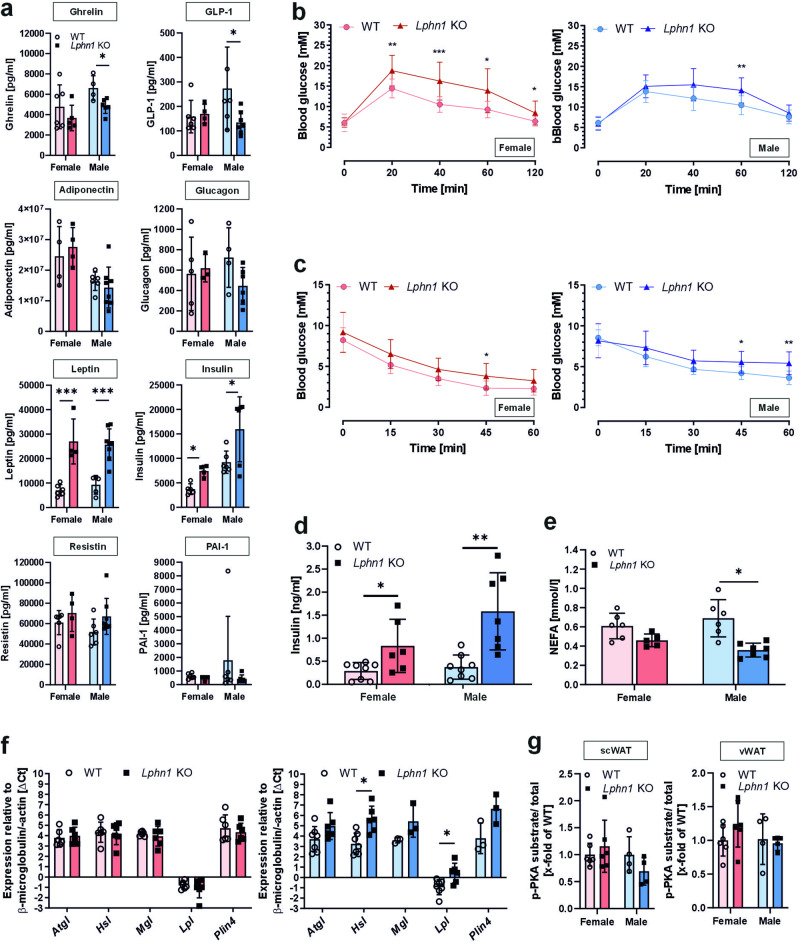
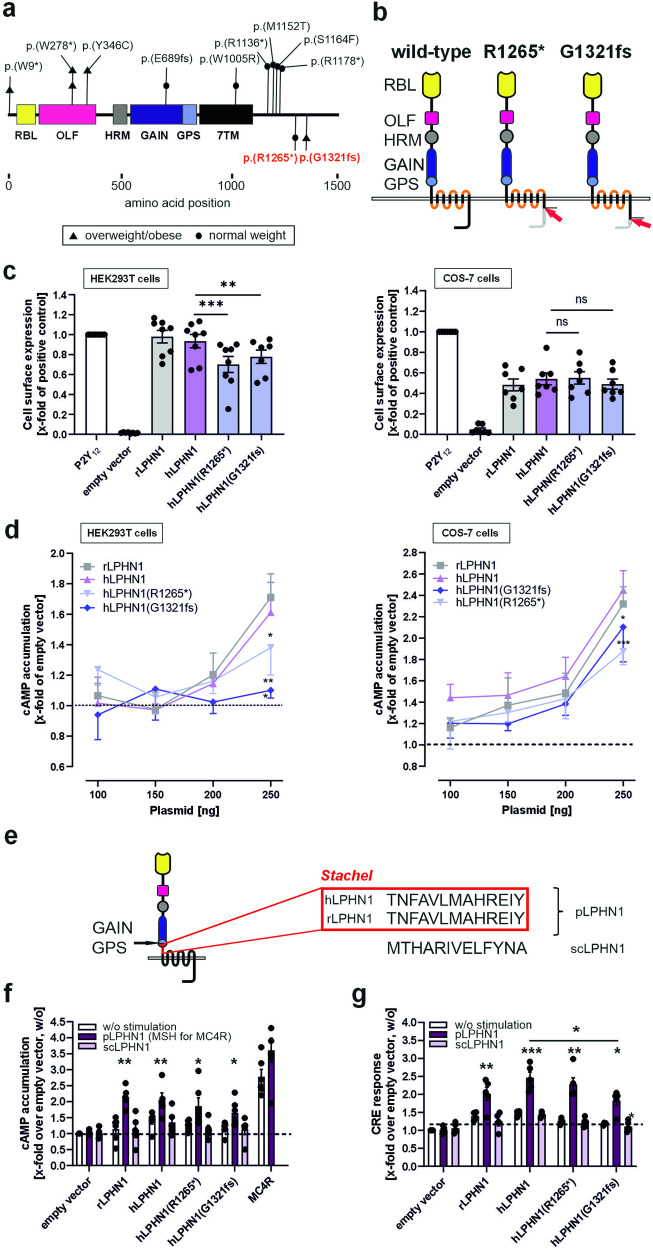
Obesity is one of the diseases with severe health consequences and rapidly increasing worldwide prevalence. Understanding the complex network of food intake and energy balance regulation is an essential prerequisite for pharmacological intervention with obesity. G protein-coupled receptors (GPCRs) are among the main modulators of metabolism and energy balance. They, for instance, regulate appetite and satiety in certain hypothalamic neurons, as well as glucose and lipid metabolism and hormone secretion from adipocytes. Mutations in some GPCRs, such as the melanocortin receptor type 4 (MC4R), have been associated with early-onset obesity. Here, we identified the adhesion GPCR latrophilin 1 (ADGRL1/LPHN1) as a member of the regulating network governing food intake and the maintenance of energy balance. Deficiency of the highly conserved receptor in mice results in increased food consumption and severe obesity, accompanied by dysregulation of glucose homeostasis. Consistently, we identified a partially inactivating mutation in human ADGRL1/LPHN1 in a patient suffering from obesity. Therefore, we propose that LPHN1 dysfunction is a risk factor for obesity development.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- NCD Risk Factor Collaboration (NCD-RisC). Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: a pooled analysis of 2416 population-based measurement studies in 128·9 million children, adolescents, and adults. Lancet (London, England). 390, 2627–2642 (2017). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
