

Methionine inducing carbohydrate esterase secretion of Trichoderma harzianum enhances the accessibility of substrate glycosidic bonds
- PMID: 38664812
- PMCID: PMC11046756
- DOI: 10.1186/s12934-024-02394-1
Methionine inducing carbohydrate esterase secretion of Trichoderma harzianum enhances the accessibility of substrate glycosidic bonds
Abstract
Background: The conversion of plant biomass into biochemicals is a promising way to alleviate energy shortage, which depends on efficient microbial saccharification and cellular metabolism. Trichoderma spp. have plentiful CAZymes systems that can utilize all-components of lignocellulose. Acetylation of polysaccharides causes nanostructure densification and hydrophobicity enhancement, which is an obstacle for glycoside hydrolases to hydrolyze glycosidic bonds. The improvement of deacetylation ability can effectively release the potential for polysaccharide degradation.
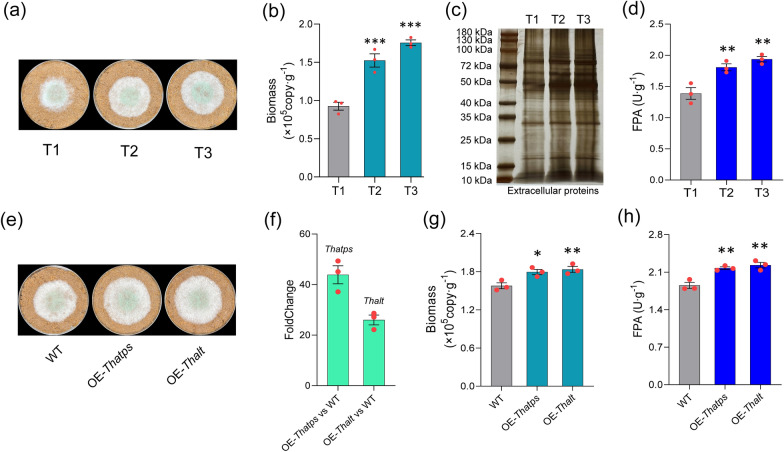
Results: Ammonium sulfate addition facilitated the deacetylation of xylan by inducing the up-regulation of multiple carbohydrate esterases (CE3/CE4/CE15/CE16) of Trichoderma harzianum. Mainly, the pathway of ammonium-sulfate's cellular assimilates inducing up-regulation of the deacetylase gene (Thce3) was revealed. The intracellular metabolite changes were revealed through metabonomic analysis. Whole genome bisulfite sequencing identified a novel differentially methylated region (DMR) that existed in the ThgsfR2 promoter, and the DMR was closely related to lignocellulolytic response. ThGsfR2 was identified as a negative regulatory factor of Thce3, and methylation in ThgsfR2 promoter released the expression of Thce3. The up-regulation of CEs facilitated the substrate deacetylation.
Conclusion: Ammonium sulfate increased the polysaccharide deacetylation capacity by inducing the up-regulation of multiple carbohydrate esterases of T. harzianum, which removed the spatial barrier of the glycosidic bond and improved hydrophilicity, and ultimately increased the accessibility of glycosidic bond to glycoside hydrolases.
Keywords: Carbohydrate esterase; Glycosidic bond accessibility; Metabolomic; Polysaccharide hydrolysis; Xylan deacetylation.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures





Similar articles
-
Comparison of fungal carbohydrate esterases of family CE16 on artificial and natural substrates.J Biotechnol. 2016 Sep 10;233:228-36. doi: 10.1016/j.jbiotec.2016.07.003. Epub 2016 Jul 18. J Biotechnol. 2016. PMID: 27439201
-
Distinct roles of carbohydrate esterase family CE16 acetyl esterases and polymer-acting acetyl xylan esterases in xylan deacetylation.J Biotechnol. 2013 Dec;168(4):684-92. doi: 10.1016/j.jbiotec.2013.10.009. Epub 2013 Oct 18. J Biotechnol. 2013. PMID: 24140638
-
A unique CE16 acetyl esterase from Podospora anserina active on polymeric xylan.Appl Microbiol Biotechnol. 2015 Dec;99(24):10515-26. doi: 10.1007/s00253-015-6934-1. Epub 2015 Sep 2. Appl Microbiol Biotechnol. 2015. PMID: 26329850
-
Microbial carbohydrate esterases deacetylating plant polysaccharides.Biotechnol Adv. 2012 Nov-Dec;30(6):1575-88. doi: 10.1016/j.biotechadv.2012.04.010. Epub 2012 May 9. Biotechnol Adv. 2012. PMID: 22580218 Review.
-
Microbial xylanolytic carbohydrate esterases.Essays Biochem. 2023 Apr 18;67(3):479-491. doi: 10.1042/EBC20220129. Essays Biochem. 2023. PMID: 36468678 Review.
Cited by
-
Toxicological and Functional Assessment of Minicell-Encapsulated dsRNA on Biocontrol Agents in Agriculture.ACS Environ Au. 2025 Jun 17;5(4):427-441. doi: 10.1021/acsenvironau.5c00067. eCollection 2025 Jul 16. ACS Environ Au. 2025. PMID: 40687506 Free PMC article.
References
-
- Xia YW, Wang JF, Guo CX, Xu HH, Wang W, Yang MZ, Shen QR, Zhang RF, Miao YZ. Exploring the multi-level regulation of lignocellulases in the filamentous fungus Trichoderma guizhouense NJAU4742 from an omics perspective. Microb Cell Fact. 2022;21:144. doi: 10.1186/s12934-022-01869-3. - DOI - PMC - PubMed
-
- Sun X, Zhang X, Huang H, Wang Y, Tu T, Bai Y, Wang Y, Zhang J, Luo H, Yao B. Engineering the cbh1 promoter of Trichoderma reesei for enhanced protein production by replacing the binding sites of a transcription repressor ACE1 to those of the activators. J Agric Food Chem. 2020;68:1337–1346. doi: 10.1021/acs.jafc.9b05452. - DOI - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
