

Engineering of Unspecific Peroxygenases Using a Superfolder-Green-Fluorescent-Protein-Mediated Secretion System in Escherichia coli
- PMID: 38665664
- PMCID: PMC11040664
- DOI: 10.1021/jacsau.4c00129
Engineering of Unspecific Peroxygenases Using a Superfolder-Green-Fluorescent-Protein-Mediated Secretion System in Escherichia coli
Abstract
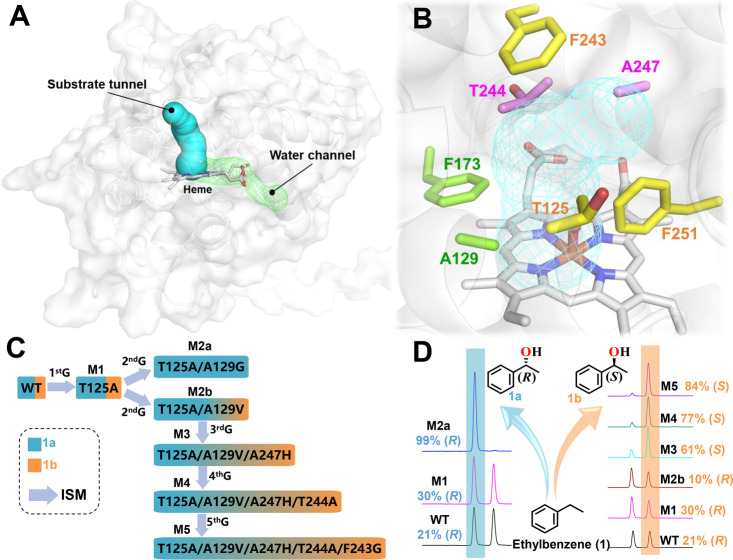
Unspecific peroxygenases (UPOs), secreted by fungi, demonstrate versatility in catalyzing challenging selective oxyfunctionalizations. However, the number of peroxygenases and corresponding variants with tailored selectivity for a broader substrate scope is still limited due to the lack of efficient engineering strategies. In this study, a new unspecific peroxygenase from Coprinopsis marcescibilis (CmaUPO) is identified and characterized. To enhance or reverse the enantioselectivity of wildtype (WT) CmaUPO catalyzed asymmetric hydroxylation of ethylbenzene, CmaUPO was engineered using an efficient superfolder-green-fluorescent-protein (sfGFP)-mediated secretion system in Escherichia coli. Iterative saturation mutagenesis (ISM) was used to target the residual sites lining the substrate tunnel, resulting in two variants: T125A/A129G and T125A/A129V/A247H/T244A/F243G. The two variants greatly improved the enantioselectivities [21% ee (R) for WT], generating the (R)-1-phenylethanol or (S)-1-phenylethanol as the main product with 99% ee (R) and 84% ee (S), respectively. The sfGFP-mediated secretion system in E. coli demonstrates applicability for different UPOs (AaeUPO, CciUPO, and PabUPO-I). Therefore, this developed system provides a robust platform for heterologous expression and enzyme engineering of UPOs, indicating great potential for their sustainable and efficient applications in various chemical transformations.
© 2024 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures





References
-
- Hofrichter M.; Kellner H.; Herzog R.; Karich A.; Liers C.; Scheibner K.; Kimani V.; Ullrich R.. Fungal peroxygenases: a phylogenetically old superfamily of heme enzymes with promiscuity for oxygen transfer reactions. In Grand Challenges in Fungal Biotechnology; Springer International Publishing, 2020; pp 369–403.10.1007/978-3-030-29541-7_14. - DOI
-
- Bormann S.; Gomez Baraibar A.; Ni Y.; Holtmann D.; Hollmann F. Specific oxyfunctionalisations catalysed by peroxygenases: opportunities, challenges and solutions. Catal. Sci. Technol. 2015, 5, 2038–2052. 10.1039/c4cy01477d. - DOI
LinkOut - more resources
Full Text Sources