

The Conventional and Breakthrough Tool for the Study of L-Glutamate Transporters
- PMID: 38668105
- PMCID: PMC11052088
- DOI: 10.3390/membranes14040077
The Conventional and Breakthrough Tool for the Study of L-Glutamate Transporters
Abstract
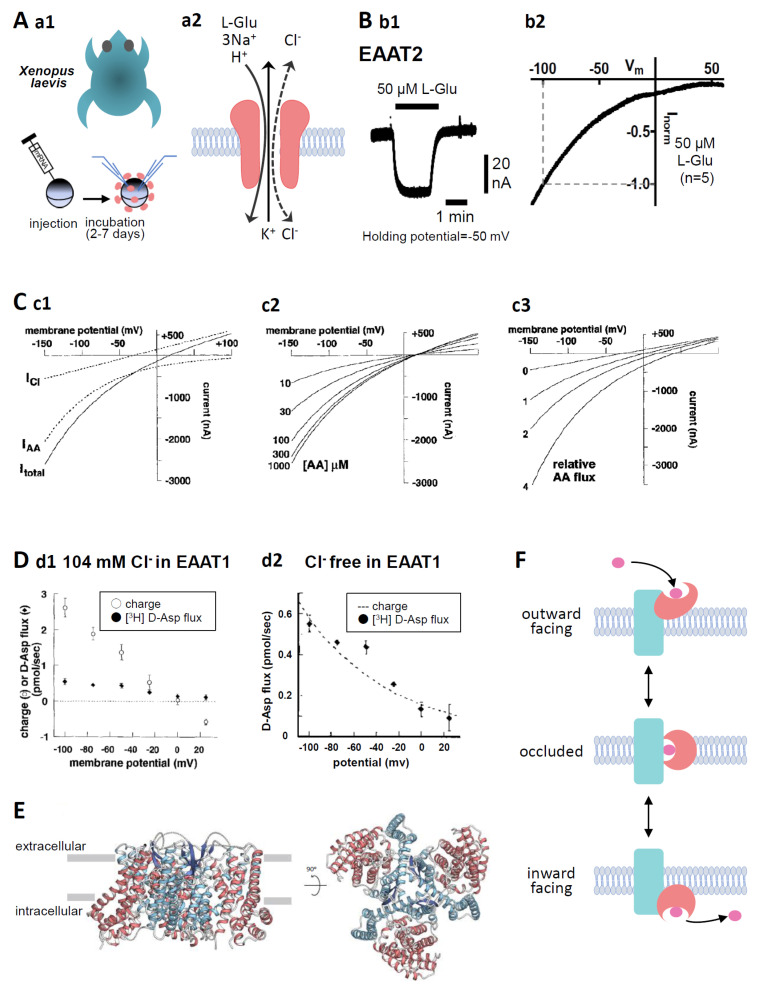
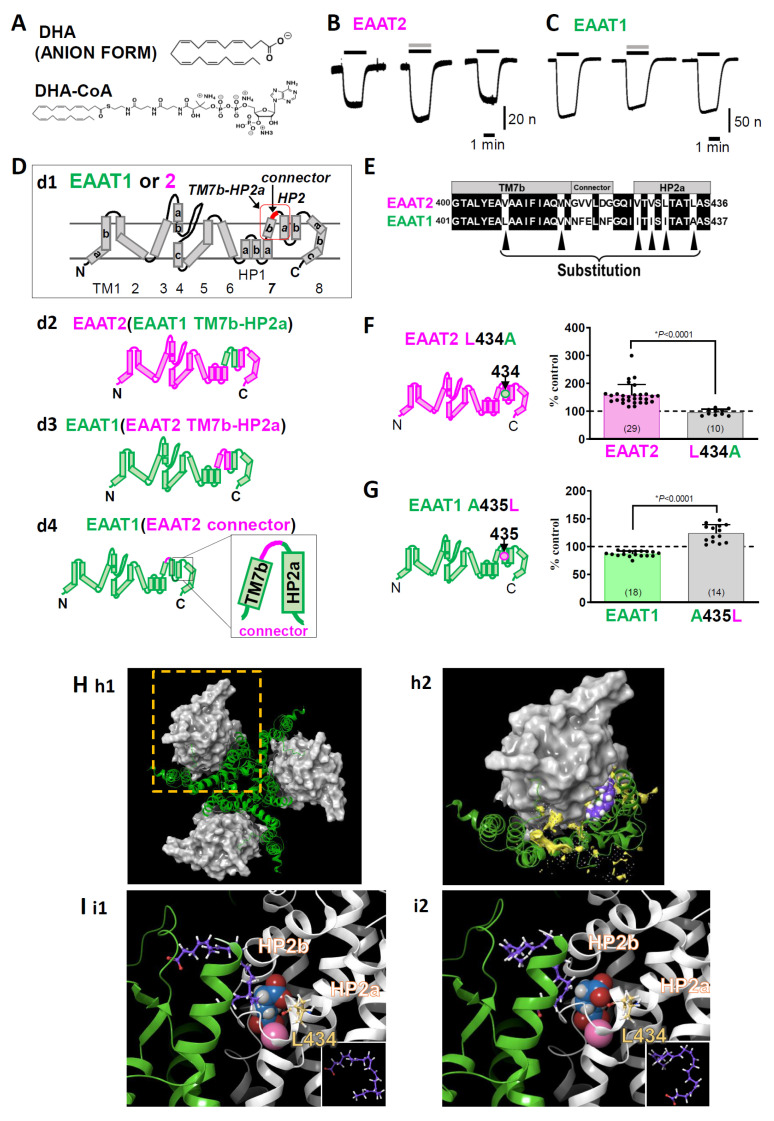
In our recent report, we clarified the direct interaction between the excitatory amino acid transporter (EAAT) 1/2 and polyunsaturated fatty acids (PUFAs) by applying electrophysiological and molecular biological techniques to Xenopus oocytes. Xenopus oocytes have a long history of use in the scientific field, but they are still attractive experimental systems for neuropharmacological studies. We will therefore summarize the pharmacological significance, advantages (especially in the study of EAAT2), and experimental techniques that can be applied to Xenopus oocytes; our new findings concerning L-glutamate (L-Glu) transporters and PUFAs; and the significant outcomes of our data. The data obtained from electrophysiological and molecular biological studies of Xenopus oocytes have provided us with further important questions, such as whether or not some PUFAs can modulate EAATs as allosteric modulators and to what extent docosahexaenoic acid (DHA) affects neurotransmission and thereby affects brain functions. Xenopus oocytes have great advantages in the studies about the interactions between molecules and functional proteins, especially in the case when the expression levels of the proteins are small in cell culture systems without transfections. These are also proper to study the mechanisms underlying the interactions. Based on the data collected in Xenopus oocyte experiments, we can proceed to the next step, i.e., the physiological roles of the compounds and their significances. In the case of EAAT2, the effects on the neurotransmission should be examined by electrophysiological approach using acute brain slices. For new drug development, pharmacokinetics pharmacodynamics (PKPD) data and blood brain barrier (BBB) penetration data are also necessary. In order not to miss the promising candidate compounds at the primary stages of drug development, we should reconsider using Xenopus oocytes in the early phase of drug development.
Keywords: EAAT2; Xenopus oocyte; excitotoxicity; glutamate transporter; overexpression; two-electrode voltage clamp (TEVC).
Conflict of interest statement
The authors declare that they have no conflicts of interest regarding the contents of this article.
Figures
Similar articles
-
Leucine 434 is essential for docosahexaenoic acid-induced augmentation of L-glutamate transporter current.J Biol Chem. 2023 Jan;299(1):102793. doi: 10.1016/j.jbc.2022.102793. Epub 2022 Dec 9. J Biol Chem. 2023. PMID: 36509140 Free PMC article.
-
Caveolin-1 Sensitivity of Excitatory Amino Acid Transporters EAAT1, EAAT2, EAAT3, and EAAT4.J Membr Biol. 2016 Jun;249(3):239-49. doi: 10.1007/s00232-015-9863-0. Epub 2015 Dec 21. J Membr Biol. 2016. PMID: 26690923
-
Regulation of the glutamate transporters by JAK2.Cell Physiol Biochem. 2011;28(4):693-702. doi: 10.1159/000335763. Epub 2011 Dec 14. Cell Physiol Biochem. 2011. PMID: 22178881
-
Endogenous amino acid transport systems and expression of mammalian amino acid transport proteins in Xenopus oocytes.Biochim Biophys Acta. 1993 Oct 29;1154(2):157-72. doi: 10.1016/0304-4157(93)90009-d. Biochim Biophys Acta. 1993. PMID: 8218336 Review.
-
Current approaches to enhance glutamate transporter function and expression.J Neurochem. 2015 Sep;134(6):982-1007. doi: 10.1111/jnc.13200. Epub 2015 Aug 14. J Neurochem. 2015. PMID: 26096891 Review.
Cited by
-
Brain pericytes upregulate glutamate uptake by astrocytes in vitro through sodium-dependent transporter.IBRO Neurosci Rep. 2025 Jun 1;19:54-61. doi: 10.1016/j.ibneur.2025.05.017. eCollection 2025 Dec. IBRO Neurosci Rep. 2025. PMID: 40535312 Free PMC article.
-
Free fatty acids inhibit an ion-coupled membrane transporter by dissipating the ion gradient.J Biol Chem. 2024 Dec;300(12):107955. doi: 10.1016/j.jbc.2024.107955. Epub 2024 Nov 2. J Biol Chem. 2024. PMID: 39491650 Free PMC article.
References
Publication types
Grants and funding
- a Research Grant on Regulatory Harmonization and Evaluation of Pharmaceuticals, Medical Devices, Regenerative and Cellular Therapy Products, Gene Therapy Products, and Cosmetics from AMED, Japan/Japan Agency for Medical Research and Development
- KAKENHI 18700373, 21700422, 17K08330/Ministry of Education, Culture, Sports, Science and Technology
- a Grant for the Program for Promotion of Fundamental Studies in Health Sciences of NIBIO/National Institute of Biomedical Innovation, Health and Nutrition
- a grant for Research on Risks of Chemicals, a Labor Science Research Grant for Research on New Drug Development/MHLW
- a Grant-in-Aid from Hoansha Foundation/Hoansha Foundation
LinkOut - more resources
Full Text Sources
