

Eliciting a single amino acid change by vaccination generates antibody protection against group 1 and group 2 influenza A viruses
- PMID: 38670113
- PMCID: PMC11096021
- DOI: 10.1016/j.immuni.2024.03.022
Eliciting a single amino acid change by vaccination generates antibody protection against group 1 and group 2 influenza A viruses
Abstract
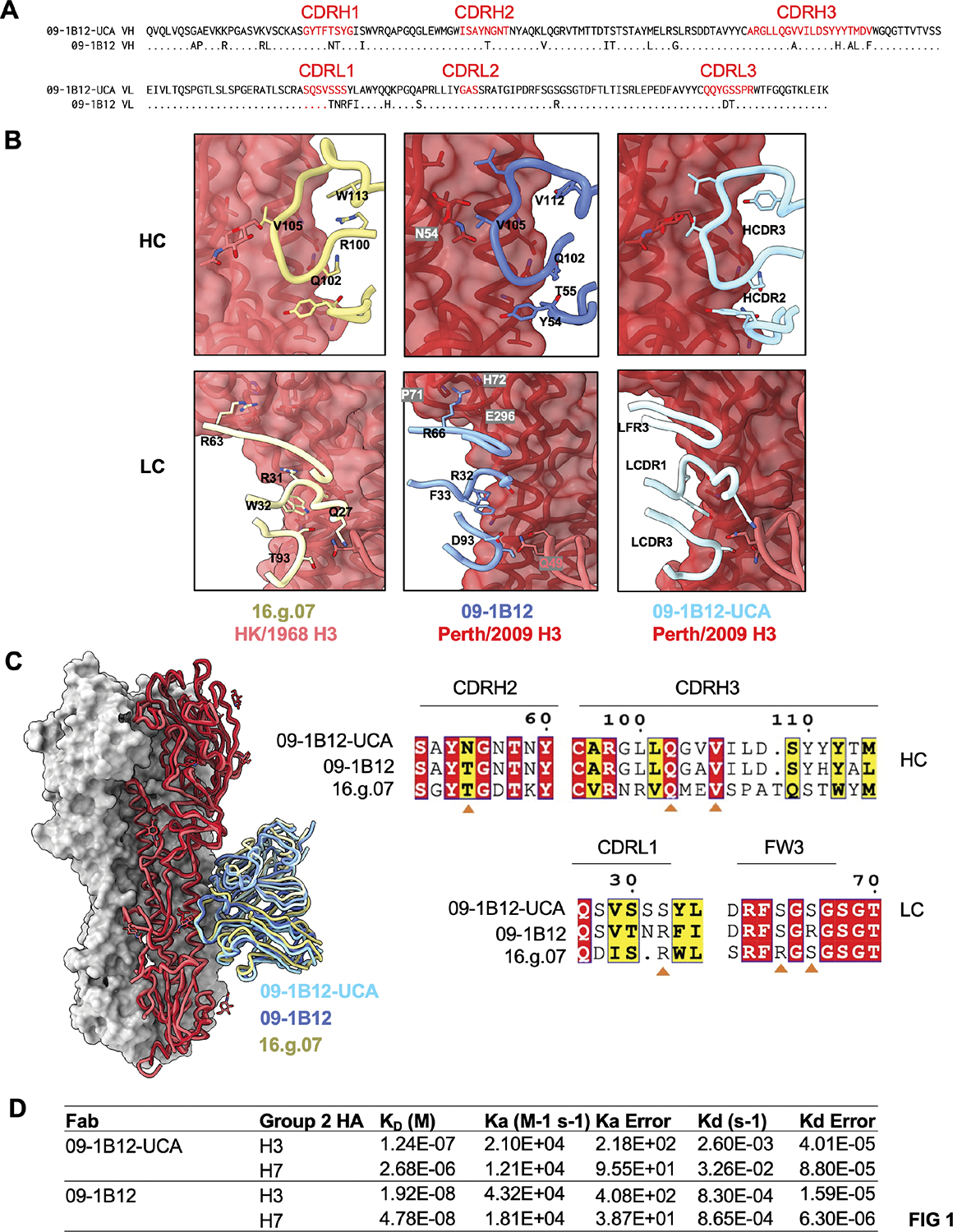
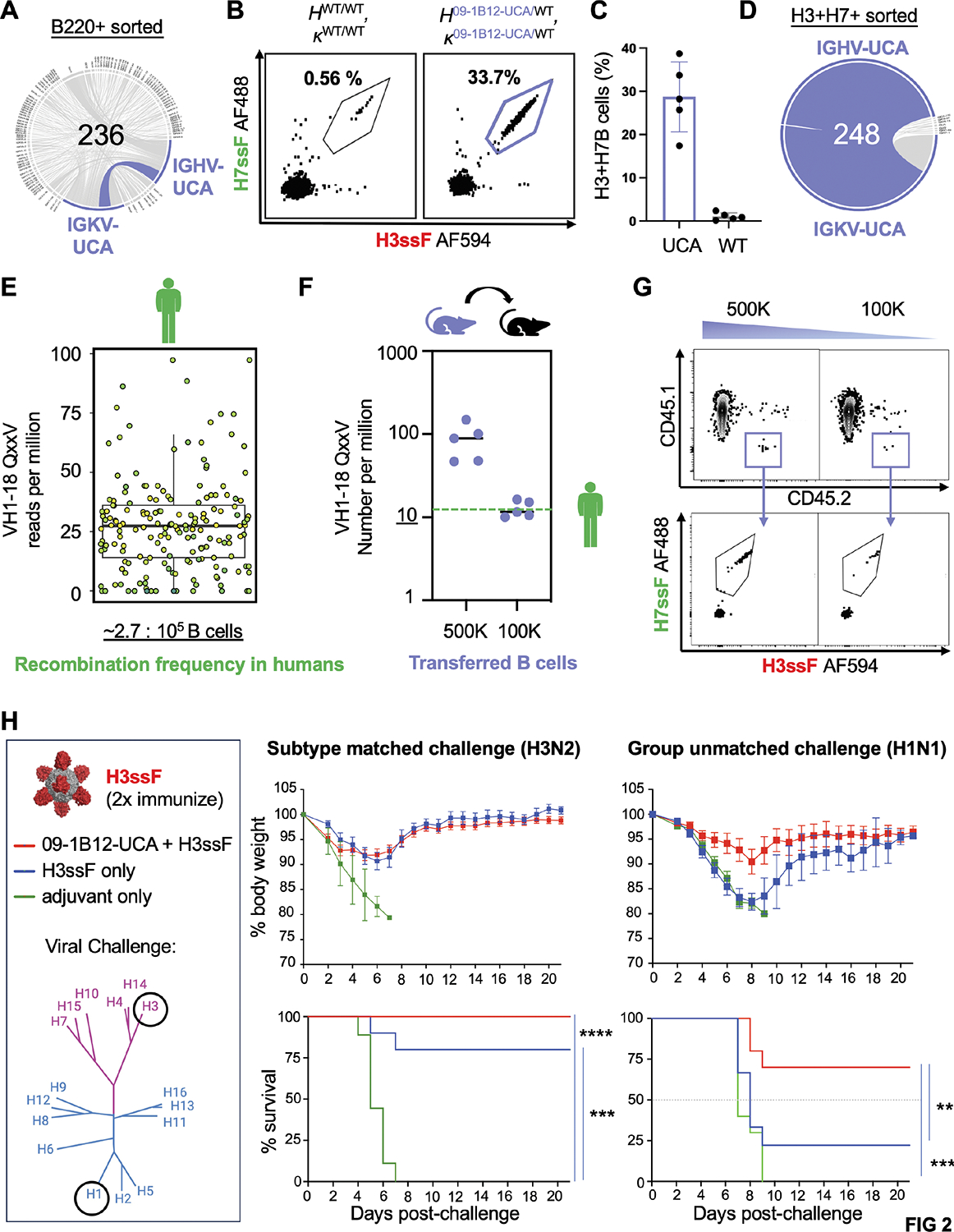
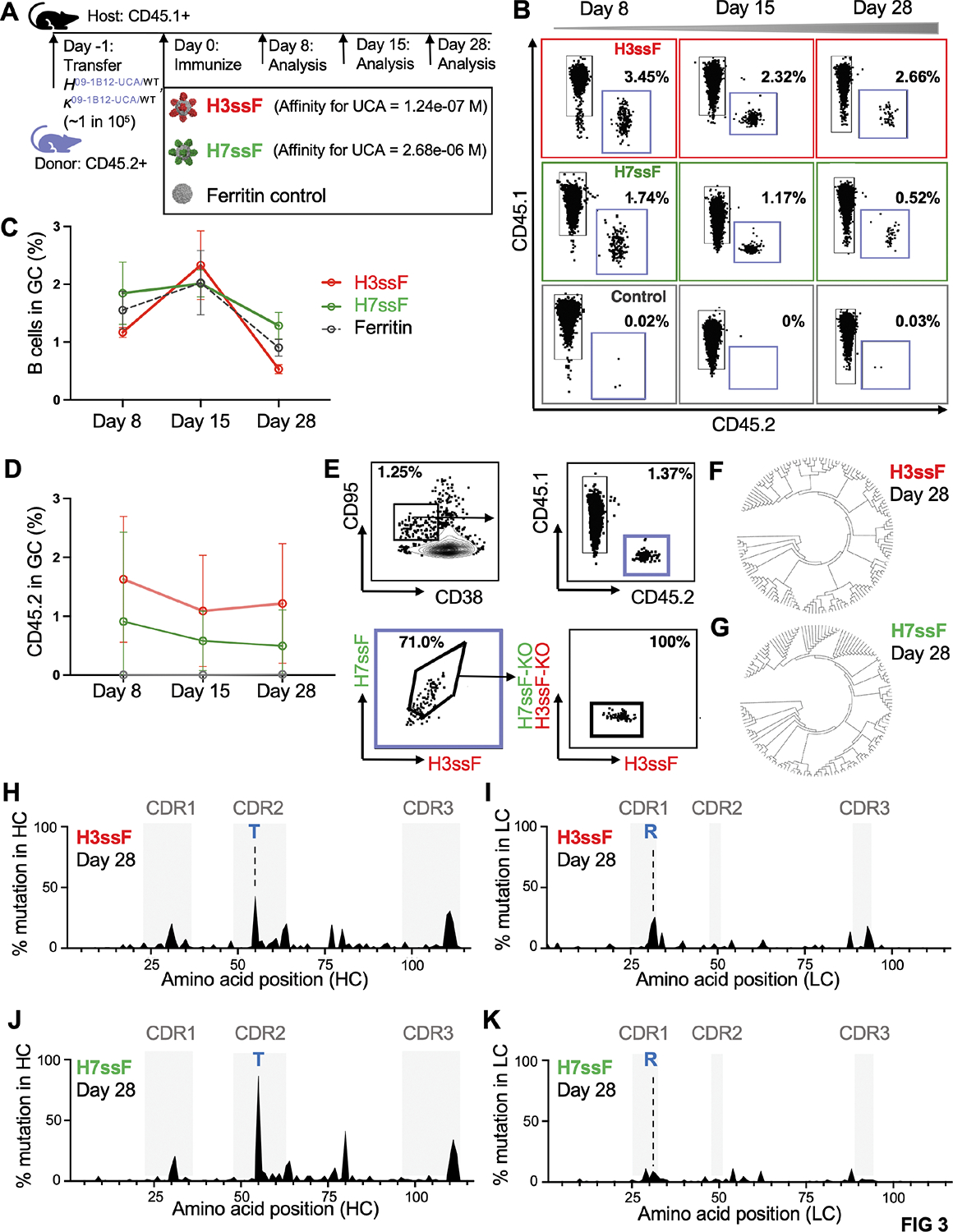
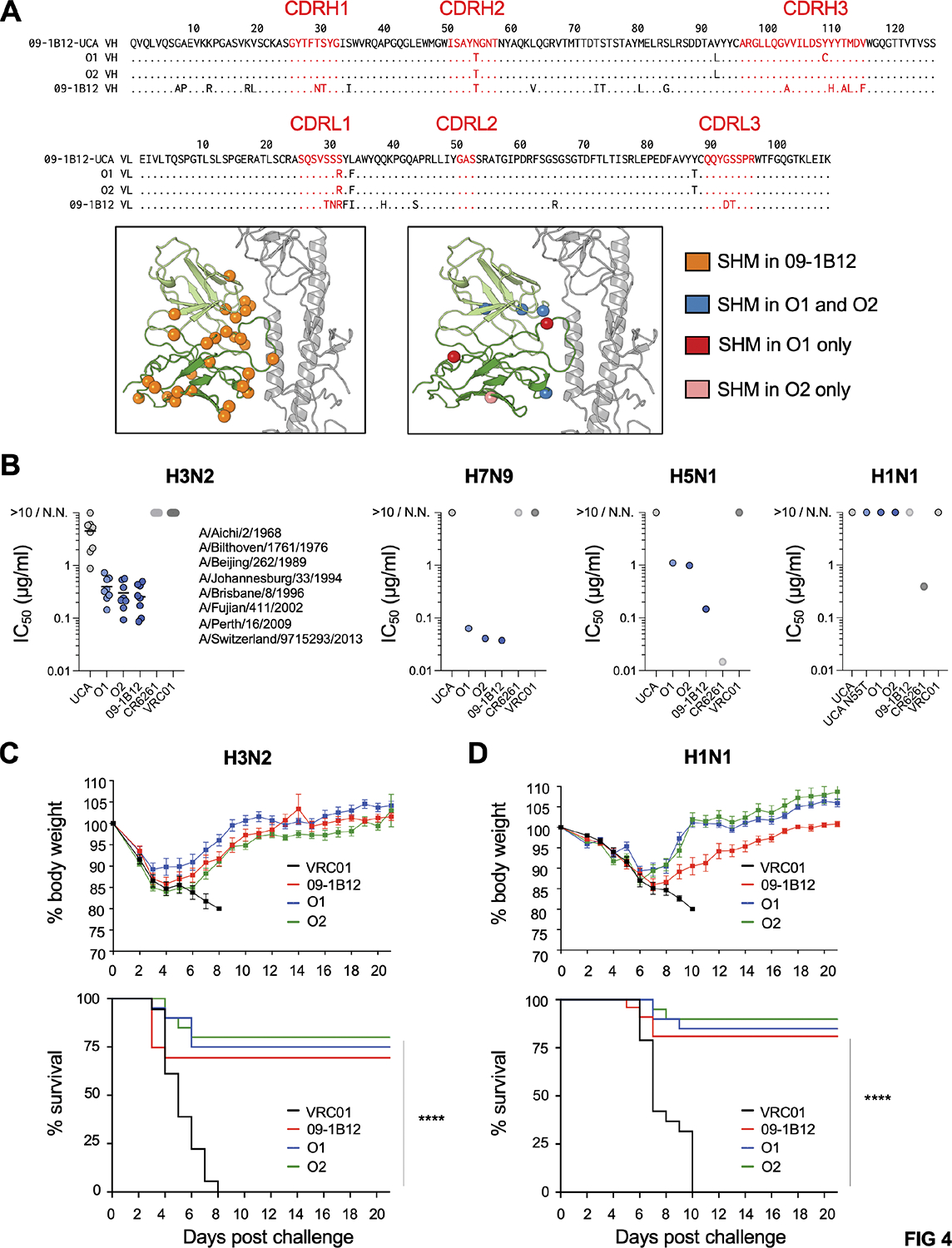
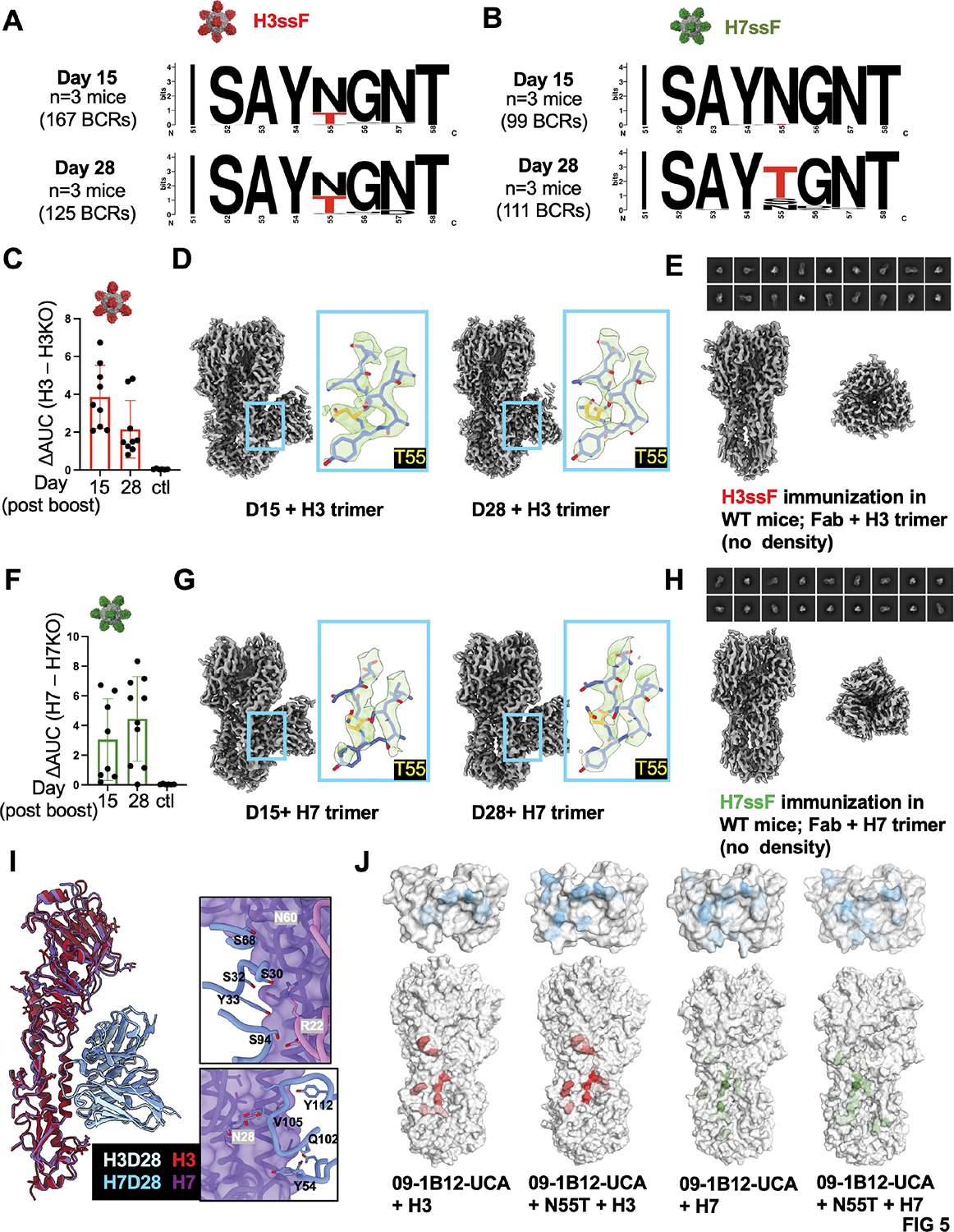
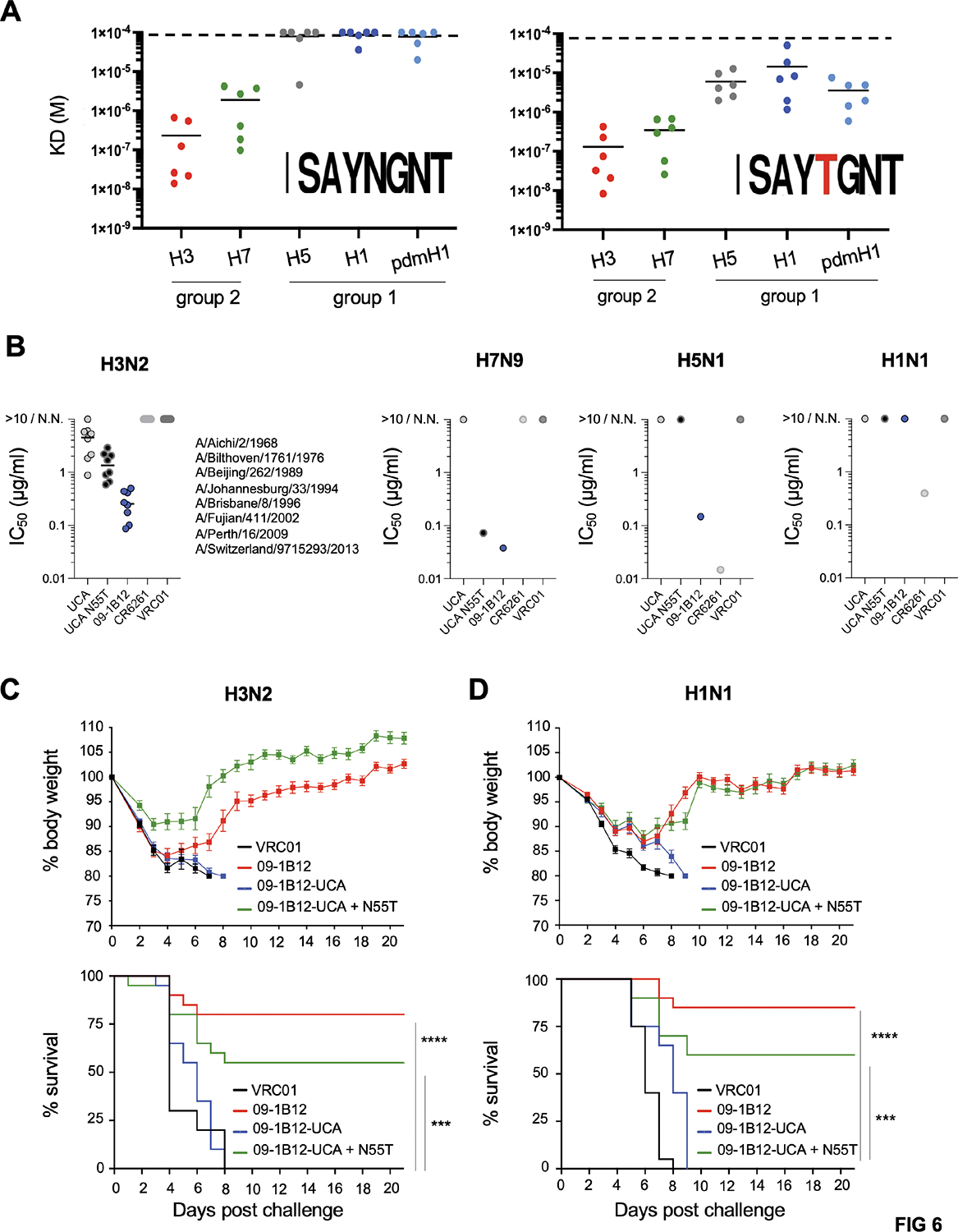
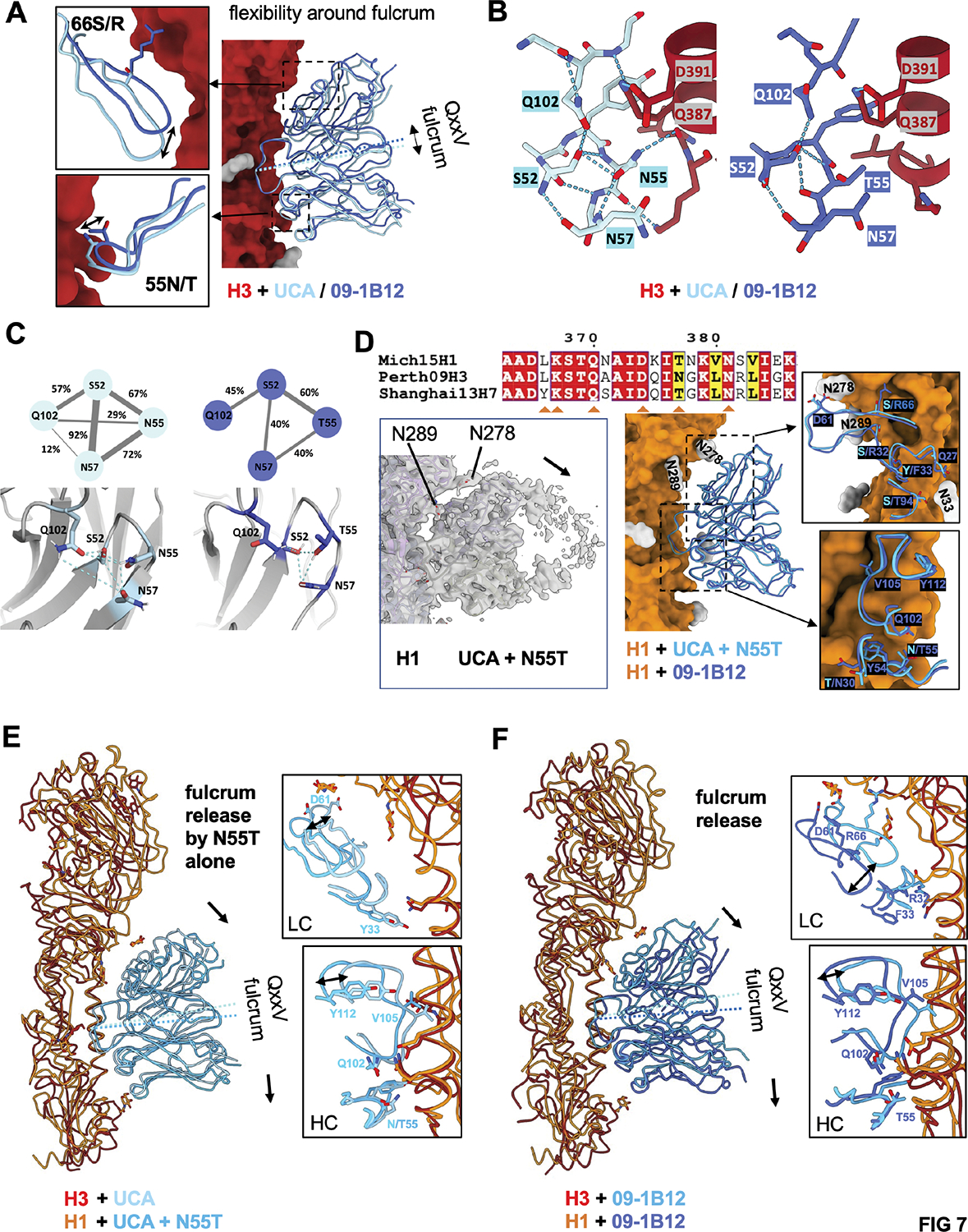
Broadly neutralizing antibodies (bnAbs) targeting the hemagglutinin (HA) stem of influenza A viruses (IAVs) tend to be effective against either group 1 or group 2 viral diversity. In rarer cases, intergroup protective bnAbs can be generated by human antibody paratopes that accommodate the conserved glycan differences between the group 1 and group 2 stems. We applied germline-engaging nanoparticle immunogens to elicit a class of cross-group bnAbs from physiological precursor frequency within a humanized mouse model. Cross-group protection depended on the presence of the human bnAb precursors within the B cell repertoire, and the vaccine-expanded antibodies enriched for an N55T substitution in the CDRH2 loop, a hallmark of the bnAb class. Structurally, this single mutation introduced a flexible fulcrum to accommodate glycosylation differences and could alone enable cross-group protection. Thus, broad IAV immunity can be expanded from the germline repertoire via minimal antigenic input and an exceptionally simple antibody development pathway.
Keywords: human antibody repertoire; immunogen; influenza virus; somatic hypermutation; universal; vaccine.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests D.L. reports SAB membership for Metaphore Bio (a Flagship company) and consultancy relationships with Tendel Therapies and Lattice Therapeutics Inc. F.D.B. has consultancy relationships with Adimab and Third Rock Ventures and is Chief Editor of The EMBO Journal. M.K. is listed as inventor of patents on vaccine immunogens used in this study filed by the U.S. Department of Health and Human Services.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
