

P2X7 receptor knockout does not alter renal function or prevent angiotensin II-induced kidney injury in F344 rats
- PMID: 38670993
- PMCID: PMC11053004
- DOI: 10.1038/s41598-024-59635-x
P2X7 receptor knockout does not alter renal function or prevent angiotensin II-induced kidney injury in F344 rats
Abstract
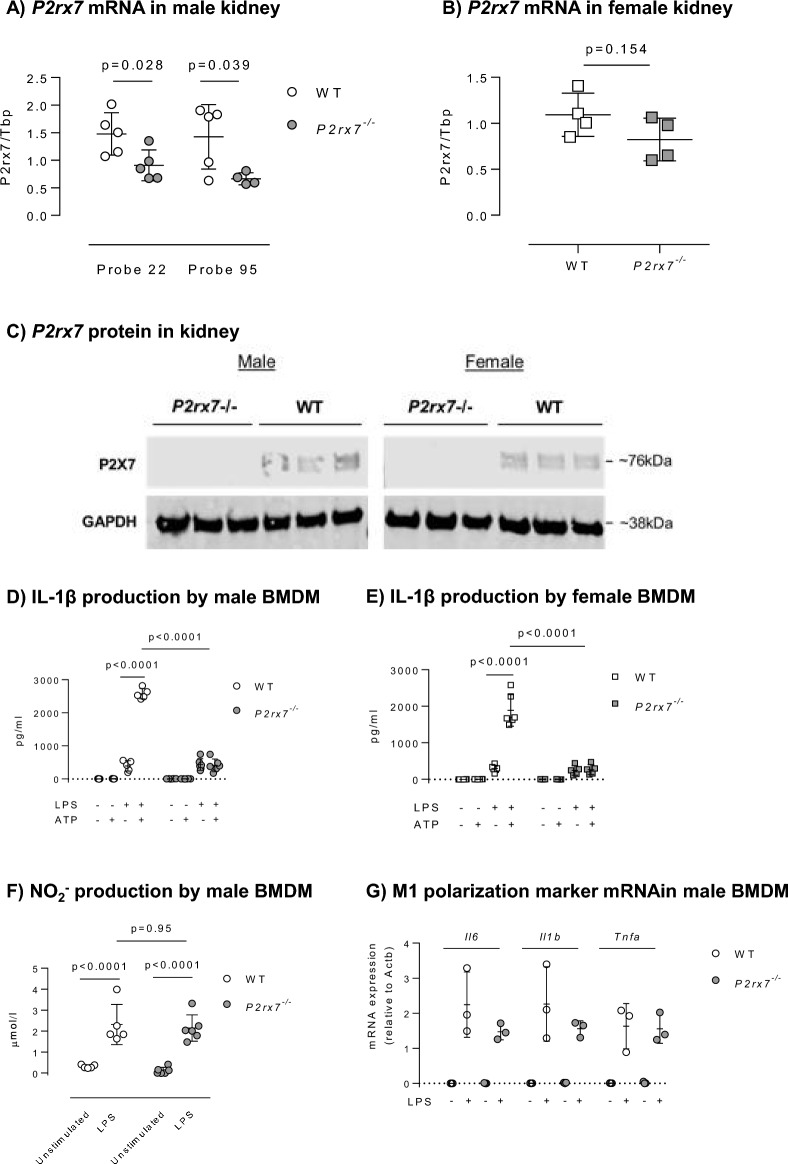
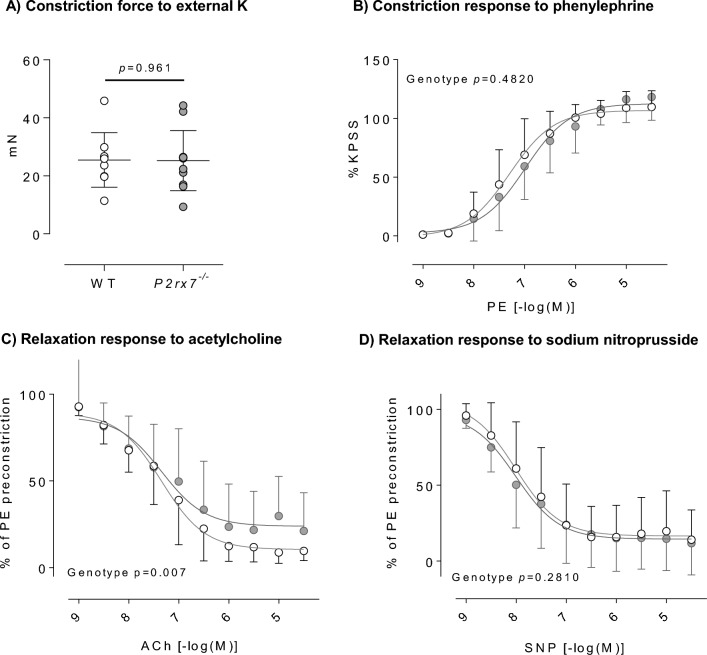
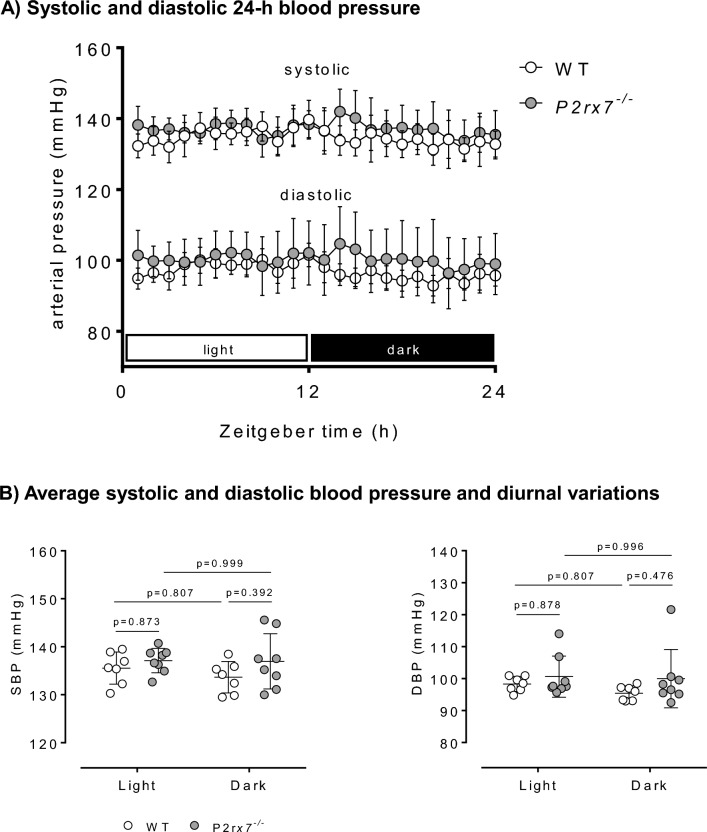
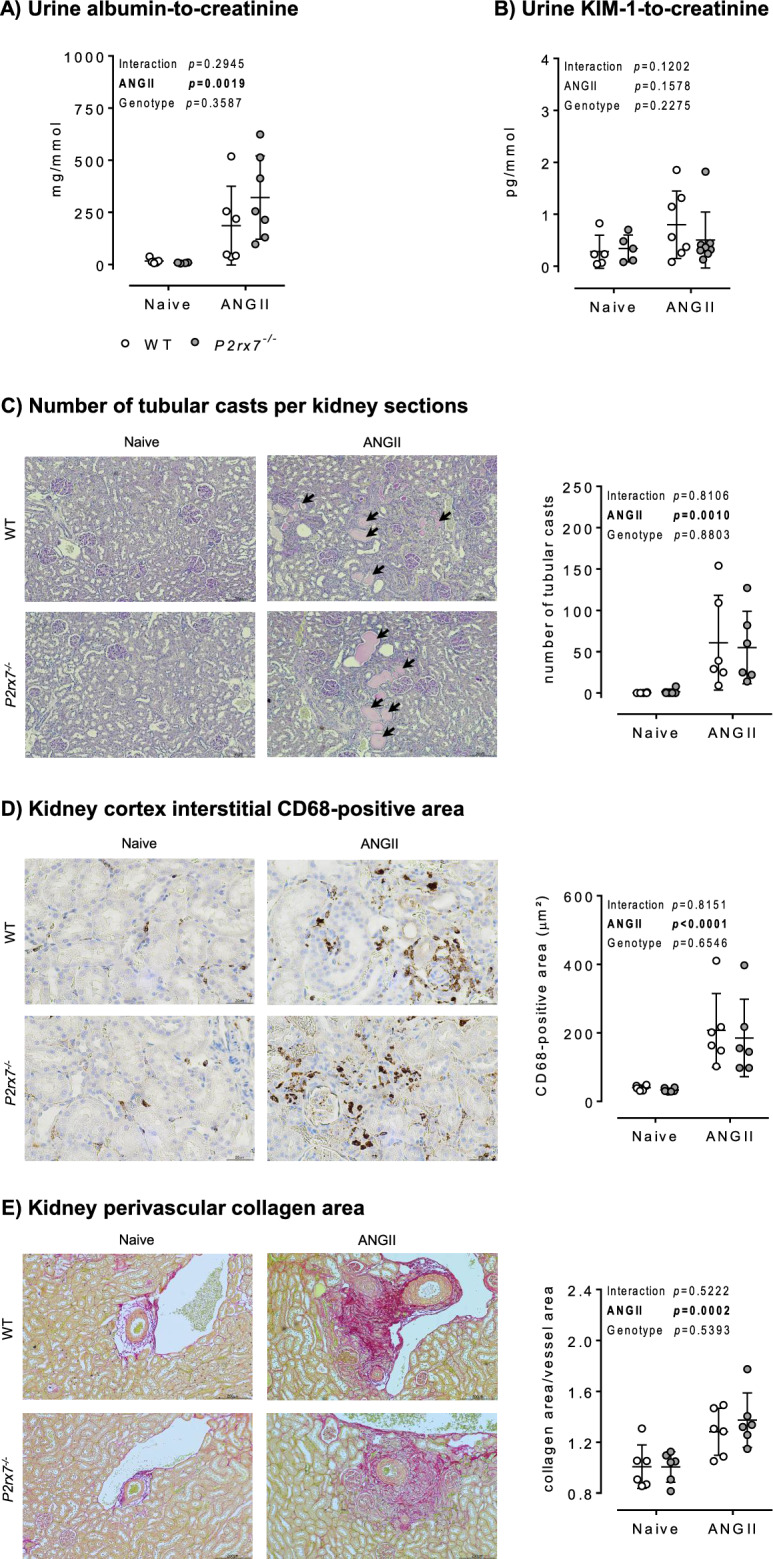
P2X7 receptors mediate immune and endothelial cell responses to extracellular ATP. Acute pharmacological blockade increases renal blood flow and filtration rate, suggesting that receptor activation promotes tonic vasoconstriction. P2X7 expression is increased in kidney disease and blockade/knockout is renoprotective. We generated a P2X7 knockout rat on F344 background, hypothesising enhanced renal blood flow and protection from angiotensin-II-induced renal injury. CRISPR/Cas9 introduced an early stop codon into exon 2 of P2rx7, abolishing P2X7 protein in kidney and reducing P2rx7 mRNA abundance by ~ 60% in bone-marrow derived macrophages. The M1 polarisation response to lipopolysaccharide was unaffected but P2X7 receptor knockout suppressed ATP-induced IL-1β release. In male knockout rats, acetylcholine-induced dilation of the renal artery ex vivo was diminished but not the response to nitroprusside. Renal function in male and female knockout rats was not different from wild-type. Finally, in male rats infused with angiotensin-II for 6 weeks, P2X7 knockout did not reduce albuminuria, tubular injury, renal macrophage accrual, and renal perivascular fibrosis. Contrary to our hypothesis, global P2X7 knockout had no impact on in vivo renal hemodynamics. Our study does not indicate a major role for P2X7 receptor activation in renal vascular injury.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests. R.I. M. has since joined AstraZeneca and may own stock or stock options.
Figures


References
-
- Burnstock, G. Purines and purinoceptors: Molecular biology overview☆. In Reference Module in Biomedical Sciences B9780128012383047413 (Elsevier, 2014). 10.1016/B978-0-12-801238-3.04741-3.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
