

Hyper-N-glycosylated SEL1L3 as auto-antigenic B-cell receptor target of primary vitreoretinal lymphomas
- PMID: 38671086
- PMCID: PMC11053041
- DOI: 10.1038/s41598-024-60169-5
Hyper-N-glycosylated SEL1L3 as auto-antigenic B-cell receptor target of primary vitreoretinal lymphomas
Abstract
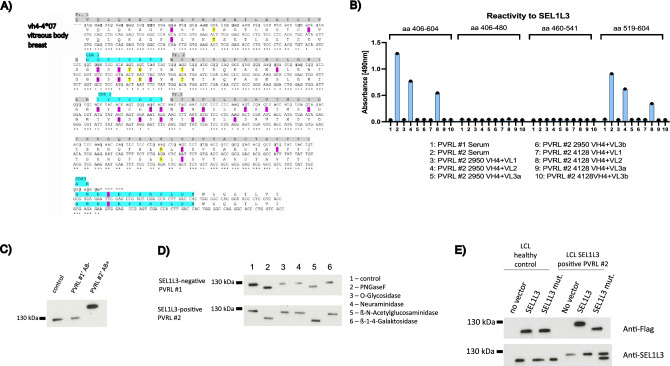
Primary vitreoretinal lymphoma (PVRL) is a rare subtype of DLBCL and can progress into primary central nervous system lymphoma (PCNSL). To investigate the role of chronic antigenic stimulation in PVRL, we cloned and expressed B-cell receptors (BCR) from PVRL patients and tested for binding against human auto-antigens. SEL1L3, a protein with multiple glycosylation sites, was identified as the BCR target in 3/20 PVRL cases. SEL1L3 induces proliferation and BCR pathway activation in aggressive lymphoma cell lines. Moreover, SEL1L3 conjugated to a toxin killed exclusively lymphoma cells with respective BCR-reactivity. Western Blot analysis indicates the occurrence of hyper-N-glycosylation of SEL1L3 at aa 527 in PVRL patients with SEL1L3-reactive BCRs. The BCR of a PVRL patient with serum antibodies against SEL1L3 was cloned from a vitreous body biopsy at diagnosis and of a systemic manifestation at relapse. VH4-04*07 was used in both lymphoma manifestations with highly conserved CDR3 regions. Both BCRs showed binding to SEL1L3, suggesting continued dependence of lymphoma cells on antigen stimulation. These results indicate an important role of antigenic stimulation by post-translationally modified auto-antigens in the genesis of PVRL. They also provide the basis for a new treatment approach targeting unique lymphoma BCRs with ultimate specificity.
Keywords: Auto-antigens; B-cell receptor antigens; Primary CNS lymphoma; Primary vitreoretinal lymphoma; SAMD14/neurabin-I; SEL1L3.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
