

The Papain-like Protease Domain of Severe Acute Respiratory Syndrome Coronavirus 2 Conjugated with Human Beta-Defensin 2 and Co1 Induces Mucosal and Systemic Immune Responses against the Virus
- PMID: 38675823
- PMCID: PMC11053661
- DOI: 10.3390/vaccines12040441
The Papain-like Protease Domain of Severe Acute Respiratory Syndrome Coronavirus 2 Conjugated with Human Beta-Defensin 2 and Co1 Induces Mucosal and Systemic Immune Responses against the Virus
Abstract
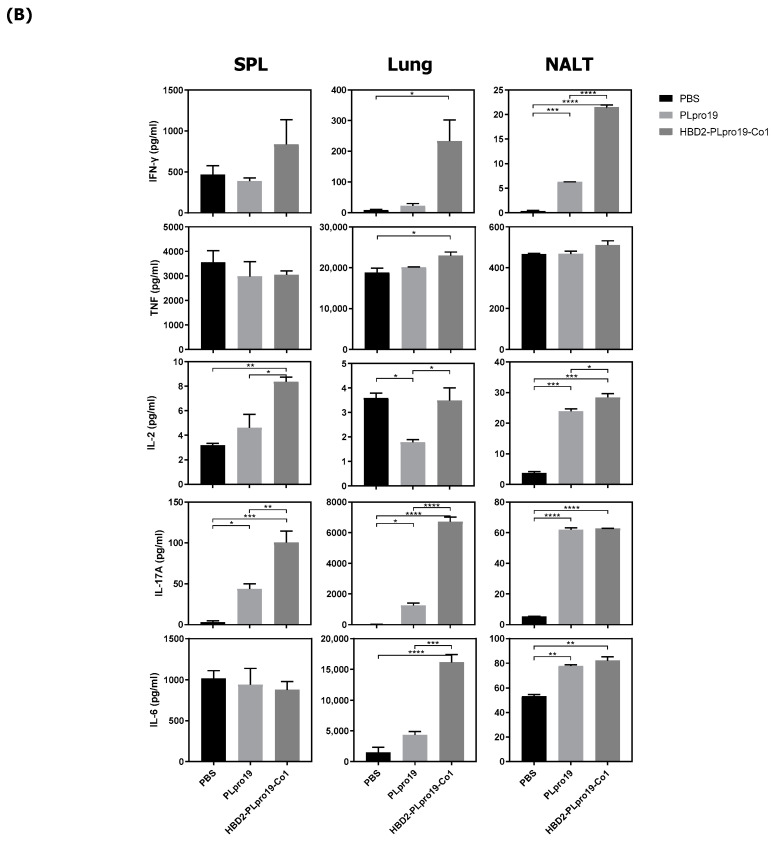
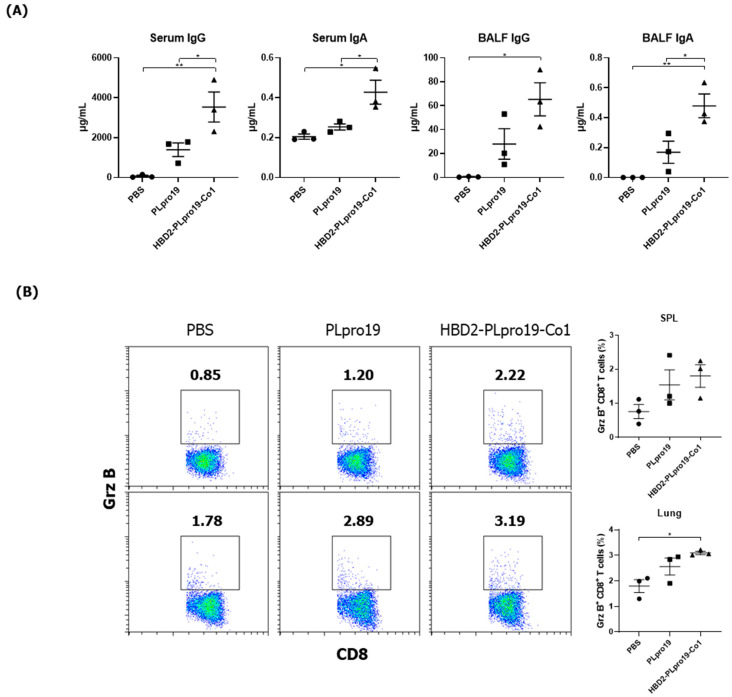
Most of the licensed vaccines against SARS-CoV-2 target spike proteins to induce viral neutralizing antibodies. However, currently prevalent SARS-CoV-2 variants contain many mutations, especially in their spike proteins. The development of vaccine antigens with conserved sequences that cross-react with variants of SARS-CoV-2 is needed to effectively defend against SARS-CoV-2 infection. Given that viral infection is initiated in the respiratory mucosa, strengthening the mucosal immune response would provide effective protection. We constructed a mucosal vaccine antigen using the papain-like protease (PLpro) domain of non-structural protein 3 of SARS-CoV-2. To potentiate the mucosal immune response, PLpro was combined with human beta-defensin 2, an antimicrobial peptide with mucosal immune adjuvant activity, and Co1, an M-cell-targeting ligand. Intranasal administration of the recombinant PLpro antigen conjugate into C57BL/6 and hACE2 knock-in (KI) mice induced antigen-specific T-cell and antibody responses with complement-dependent cytotoxic activity. Viral challenge experiments using the Wuhan and Delta strains of SARS-CoV-2 provided further evidence that immunized hACE2 KI mice were protected against viral challenge infections. Our study shows that PLpro is a useful candidate vaccine antigen against SARS-CoV-2 infection and that the inclusion of human beta-defensin 2 and Co1 in the recombinant construct may enhance the efficacy of the vaccine.
Keywords: SARS-CoV-2; adjuvant; papain-like protease; recombinant antigen; vaccine.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures










Similar articles
-
A Beta Strain-Based Spike Glycoprotein Vaccine Candidate Induces Broad Neutralization and Protection against SARS-CoV-2 Variants of Concern.Microbiol Spectr. 2023 Feb 27;11(2):e0268722. doi: 10.1128/spectrum.02687-22. Online ahead of print. Microbiol Spectr. 2023. PMID: 36847495 Free PMC article.
-
N-dihydrogalactochitosan serves as an effective mucosal adjuvant for intranasal vaccine in combination with recombinant viral proteins against respiratory infection.Acta Biomater. 2024 Feb;175:279-292. doi: 10.1016/j.actbio.2023.12.039. Epub 2023 Dec 29. Acta Biomater. 2024. PMID: 38160856
-
Intranasal administration of a single dose of MVA-based vaccine candidates against COVID-19 induced local and systemic immune responses and protects mice from a lethal SARS-CoV-2 infection.Front Immunol. 2022 Sep 12;13:995235. doi: 10.3389/fimmu.2022.995235. eCollection 2022. Front Immunol. 2022. PMID: 36172368 Free PMC article.
-
The SARS-coronavirus papain-like protease: structure, function and inhibition by designed antiviral compounds.Antiviral Res. 2015 Mar;115:21-38. doi: 10.1016/j.antiviral.2014.12.015. Epub 2014 Dec 29. Antiviral Res. 2015. PMID: 25554382 Free PMC article. Review.
-
COVID-19 Pandemic and Vaccines Update on Challenges and Resolutions.Front Cell Infect Microbiol. 2021 Sep 10;11:690621. doi: 10.3389/fcimb.2021.690621. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34568087 Free PMC article. Review.
References
-
- Chakraborty C., Bhattacharya M., Dhama K. SARS-CoV-2 vaccines, vaccine development technologies, and significant efforts in vaccine development during the pandemic: The lessons learned might help to fight against the next pandemic. Vaccines. 2023;11:682. doi: 10.3390/vaccines11030682. - DOI - PMC - PubMed
-
- Chen J., Xu W., Li L., Yi L., Jiang Y., Hao P., Xu Z., Zou W., Li P., Gao Z. Immunogenicity and protective potential of chimeric virus-like particles containing SARS-CoV-2 spike and H5N1 matrix 1 proteins. Front. Cell. Infect. Microbiol. 2022;12:967493. doi: 10.3389/fcimb.2022.967493. - DOI - PMC - PubMed
-
- Li L., Honda-Okubo Y., Huang Y., Jang H., Carlock M.A., Baldwin J., Piplani S., Bebin-Blackwell A.G., Forgacs D., Sakamoto K. Immunisation of ferrets and mice with recombinant SARS-CoV-2 spike protein formulated with Advax-SM adjuvant protects against COVID-19 infection. Vaccine. 2021;39:5940–5953. doi: 10.1016/j.vaccine.2021.07.087. - DOI - PMC - PubMed
-
- Korber B., Fischer W.M., Gnanakaran S., Yoon H., Theiler J., Abfalterer W., Hengartner N., Giorgi E.E., Bhattacharya T., Foley B. Tracking changes in SARS-CoV-2 spike: Evidence that D614G increases infectivity of the COVID-19 virus. Cell. 2020;182:812–827.e19. doi: 10.1016/j.cell.2020.06.043. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
