

Efficient gene knockout and genetic interaction screening using the in4mer CRISPR/Cas12a multiplex knockout platform
- PMID: 38678031
- PMCID: PMC11055879
- DOI: 10.1038/s41467-024-47795-3
Efficient gene knockout and genetic interaction screening using the in4mer CRISPR/Cas12a multiplex knockout platform
Abstract
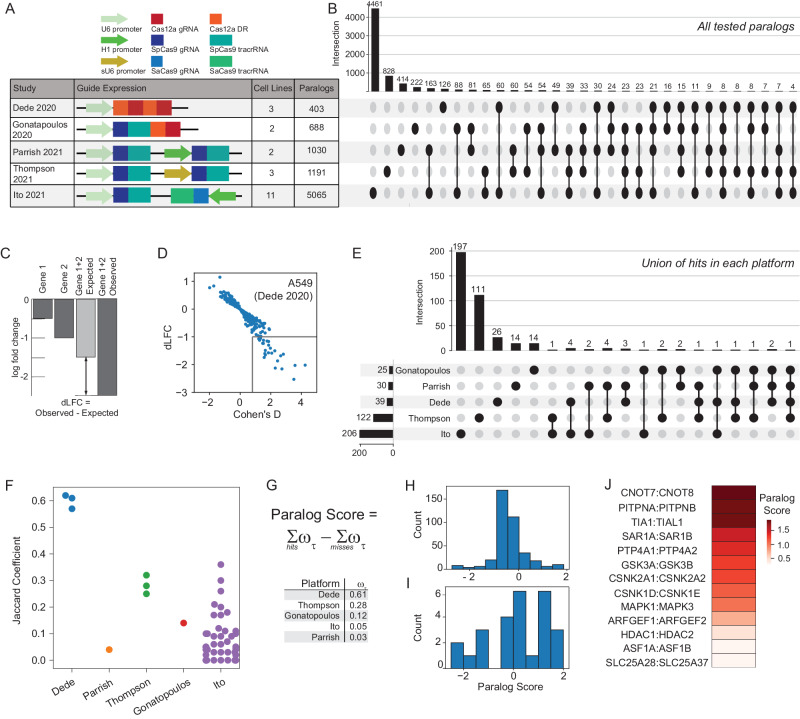
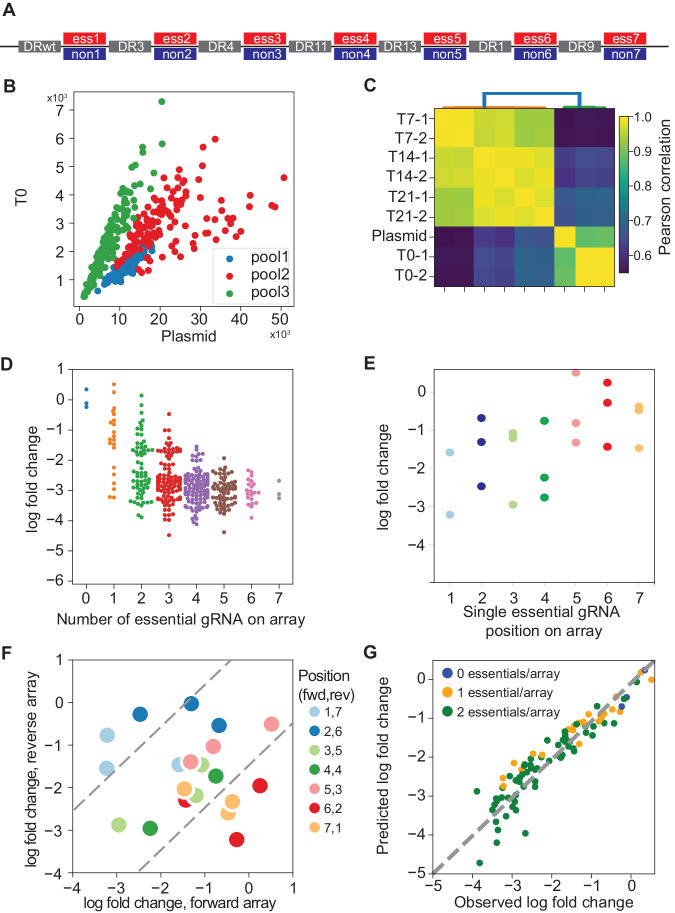
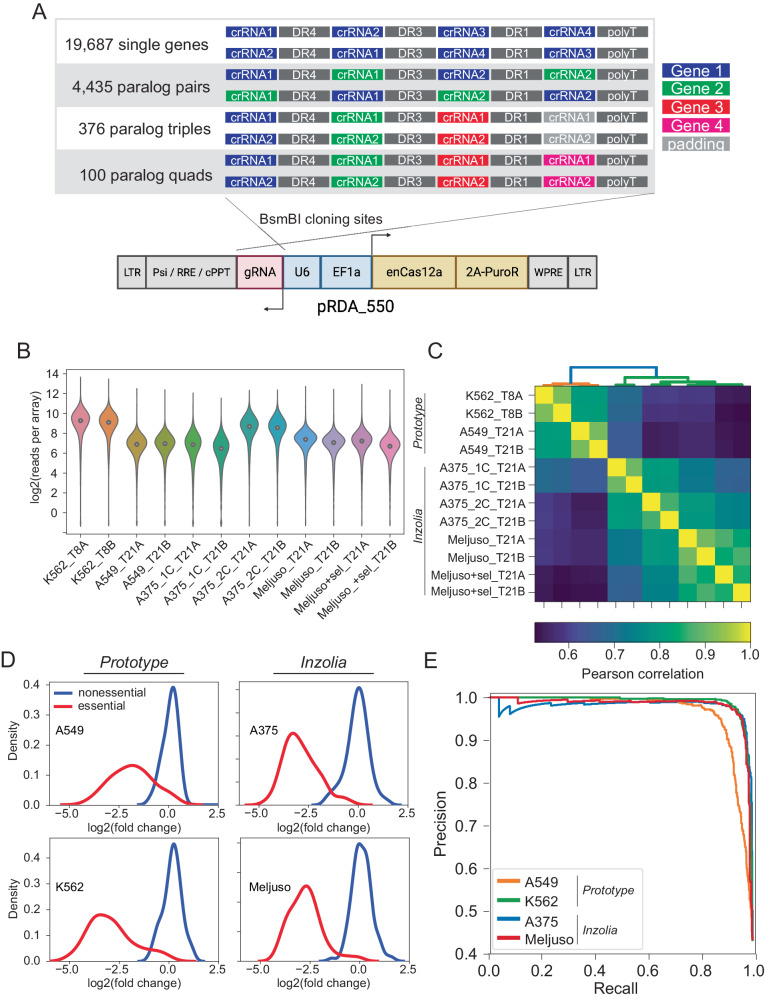
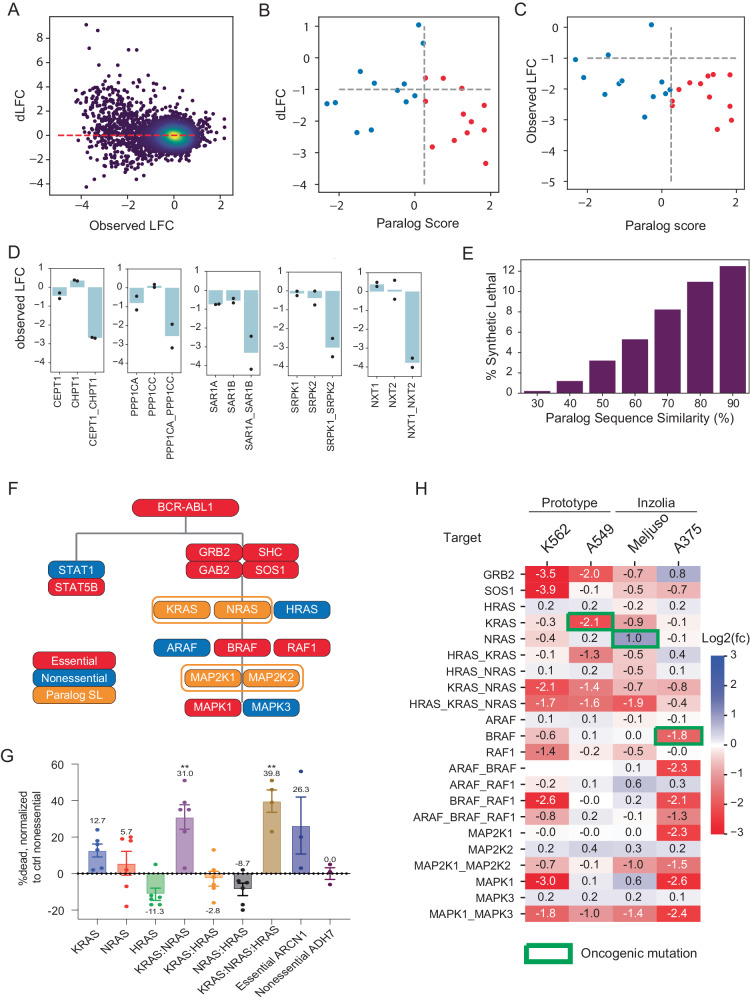
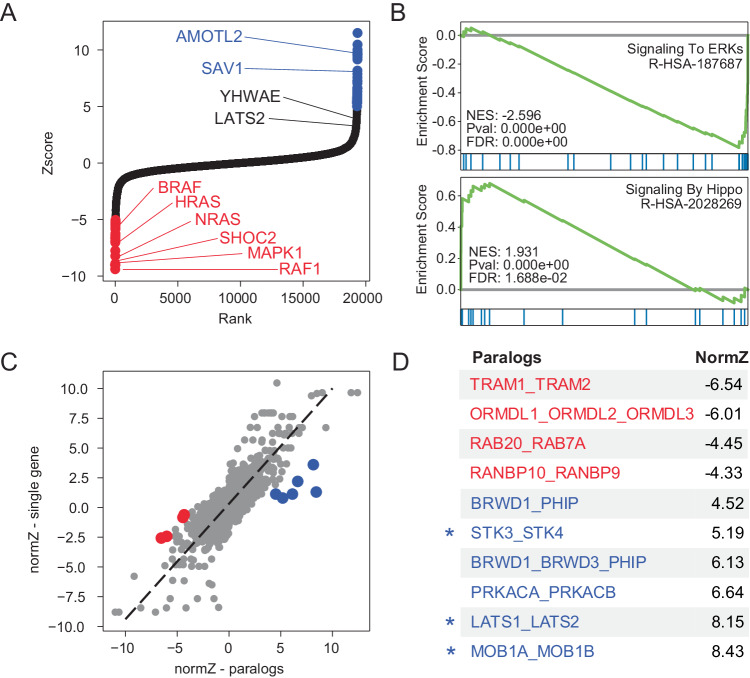
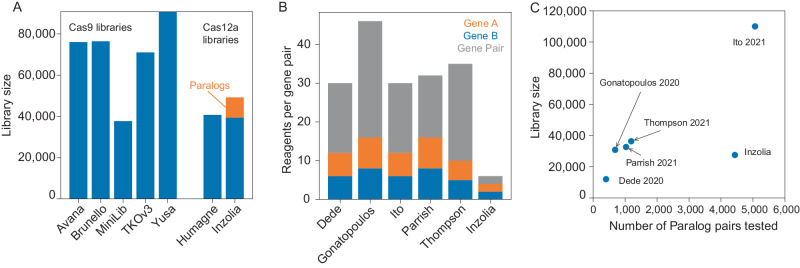
Genetic interactions mediate the emergence of phenotype from genotype, but technologies for combinatorial genetic perturbation in mammalian cells are challenging to scale. Here, we identify background-independent paralog synthetic lethals from previous CRISPR genetic interaction screens, and find that the Cas12a platform provides superior sensitivity and assay replicability. We develop the in4mer Cas12a platform that uses arrays of four independent guide RNAs targeting the same or different genes. We construct a genome-scale library, Inzolia, that is ~30% smaller than a typical CRISPR/Cas9 library while also targeting ~4000 paralog pairs. Screens in cancer cells demonstrate discrimination of core and context-dependent essential genes similar to that of CRISPR/Cas9 libraries, as well as detection of synthetic lethal and masking/buffering genetic interactions between paralogs of various family sizes. Importantly, the in4mer platform offers a fivefold reduction in library size compared to other genetic interaction methods, substantially reducing the cost and effort required for these assays.
© 2024. The Author(s).
Conflict of interest statement
J.G.D. consults for Microsoft Research, Abata Therapeutics, Servier, Maze Therapeutics, BioNTech, Sangamo, and Pfizer. J.G.D. consults for and has equity in Tango Therapeutics. J.G.D. serves as a paid scientific advisor to the Laboratory for Genomics Research, funded in part by GlaxoSmithKline. J.G.D. receives funding support from the Functional Genomics Consortium: Abbvie, Bristol Myers Squibb, Janssen, Merck, and Vir Biotechnology. J.G.D.’s interests were reviewed and are managed by the Broad Institute in accordance with its conflict of interest policies. Other authors don’t claim competing interests.
Figures
Update of
-
Efficient gene knockout and genetic interactions: the IN4MER CRISPR/Cas12a multiplex knockout platform.bioRxiv [Preprint]. 2023 Sep 5:2023.01.03.522655. doi: 10.1101/2023.01.03.522655. bioRxiv. 2023. Update in: Nat Commun. 2024 Apr 27;15(1):3577. doi: 10.1038/s41467-024-47795-3. PMID: 36712129 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
