

Psilocybin restrains activity-based anorexia in female rats by enhancing cognitive flexibility: contributions from 5-HT1A and 5-HT2A receptor mechanisms
- PMID: 38678087
- PMCID: PMC11449803
- DOI: 10.1038/s41380-024-02575-9
Psilocybin restrains activity-based anorexia in female rats by enhancing cognitive flexibility: contributions from 5-HT1A and 5-HT2A receptor mechanisms
Abstract
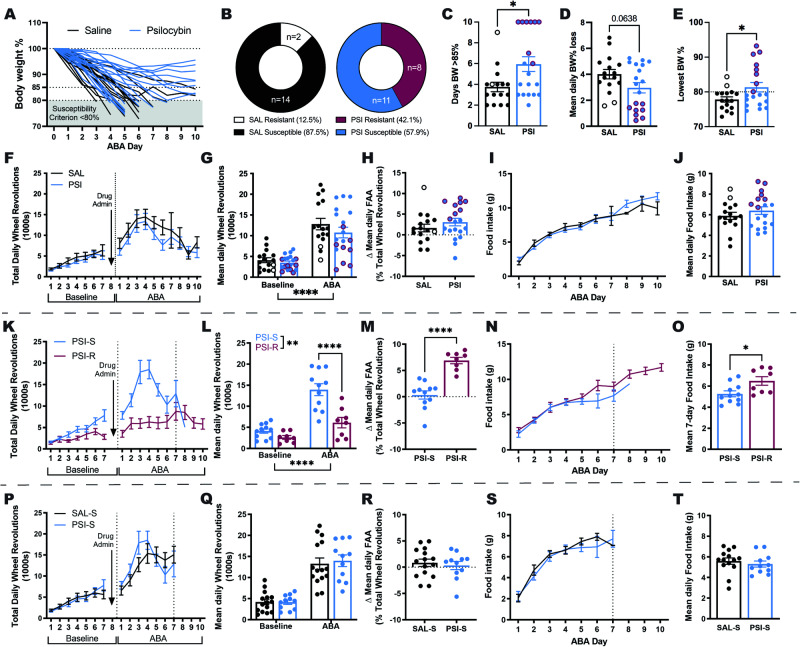
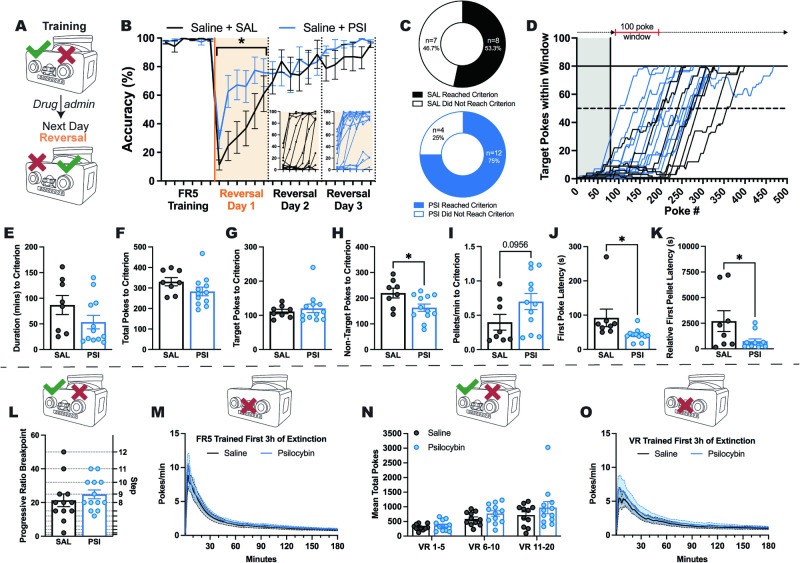
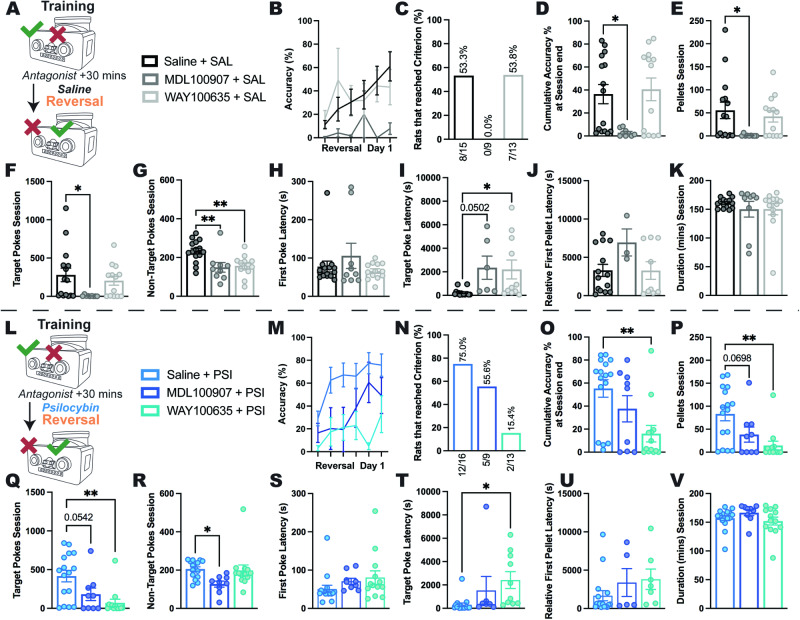
Psilocybin has shown promise for alleviating symptoms of depression and is currently in clinical trials for the treatment of anorexia nervosa (AN), a condition that is characterised by persistent cognitive inflexibility. Considering that enhanced cognitive flexibility after psilocybin treatment is reported to occur in individuals with depression, it is plausible that psilocybin could improve symptoms of AN by breaking down cognitive inflexibility. A mechanistic understanding of the actions of psilocybin is required to tailor the clinical application of psilocybin to individuals most likely to respond with positive outcomes. This can only be achieved using incisive neurobiological approaches in animal models. Here, we use the activity-based anorexia (ABA) rat model and comprehensively assess aspects of reinforcement learning to show that psilocybin (post-acutely) improves body weight maintenance in female rats and facilitates cognitive flexibility, specifically via improved adaptation to the initial reversal of reward contingencies. Further, we reveal the involvement of signalling through the serotonin (5-HT) 1 A and 5-HT2A receptor subtypes in specific aspects of learning, demonstrating that 5-HT1A antagonism negates the cognitive enhancing effects of psilocybin. Moreover, we show that psilocybin elicits a transient increase and decrease in cortical transcription of these receptors (Htr2a and Htr1a, respectively), and a further reduction in the abundance of Htr2a transcripts in rats exposed to the ABA model. Together, these findings support the hypothesis that psilocybin could ameliorate cognitive inflexibility in the context of AN and highlight a need to better understand the therapeutic mechanisms independent of 5-HT2A receptor binding.
© 2024. The Author(s).
Conflict of interest statement
CJF sits on the scientific advisory board for Octarine Bio, Copenhagen, Denmark.
Figures


Similar articles
-
Separate or inseparable? Serotonin and dopamine system interactions may underlie the therapeutic potential of psilocybin for anorexia nervosa.Physiol Behav. 2025 Sep 1;298:114957. doi: 10.1016/j.physbeh.2025.114957. Epub 2025 May 20. Physiol Behav. 2025. PMID: 40403997 Review.
-
The 5-HT2A/1A agonist psilocybin disrupts modal object completion associated with visual hallucinations.Biol Psychiatry. 2011 Mar 1;69(5):399-406. doi: 10.1016/j.biopsych.2010.10.002. Epub 2010 Dec 3. Biol Psychiatry. 2011. PMID: 21126732
-
Rapid, automated, and experimenter-free touchscreen testing reveals reciprocal interactions between cognitive flexibility and activity-based anorexia in female rats.Elife. 2023 Jun 30;12:e84961. doi: 10.7554/eLife.84961. Elife. 2023. PMID: 37387293 Free PMC article.
-
Activation of serotonin 2A receptors underlies the psilocybin-induced effects on α oscillations, N170 visual-evoked potentials, and visual hallucinations.J Neurosci. 2013 Jun 19;33(25):10544-51. doi: 10.1523/JNEUROSCI.3007-12.2013. J Neurosci. 2013. PMID: 23785166 Free PMC article. Clinical Trial.
-
Rethinking Therapeutic Strategies for Anorexia Nervosa: Insights From Psychedelic Medicine and Animal Models.Front Neurosci. 2020 Feb 4;14:43. doi: 10.3389/fnins.2020.00043. eCollection 2020. Front Neurosci. 2020. PMID: 32116500 Free PMC article. Review.
Cited by
-
Potential Differences in Psychedelic Actions Based on Biological Sex.Endocrinology. 2024 Jul 1;165(8):bqae083. doi: 10.1210/endocr/bqae083. Endocrinology. 2024. PMID: 38980913 Free PMC article. Review.
-
The Emergence of Psilocybin in Psychiatry and Neuroscience.Pharmaceuticals (Basel). 2025 Apr 9;18(4):555. doi: 10.3390/ph18040555. Pharmaceuticals (Basel). 2025. PMID: 40283990 Free PMC article. Review.
-
Unraveling psilocybin's therapeutic potential: behavioral and neuroplasticity insights in Wistar-Kyoto and Wistar male rat models of treatment-resistant depression.Psychopharmacology (Berl). 2025 Jul;242(7):1607-1625. doi: 10.1007/s00213-024-06644-3. Epub 2024 Jul 4. Psychopharmacology (Berl). 2025. PMID: 38963553 Free PMC article.
-
Psychedelic enhancement of flexible learning weeks after a single dose.bioRxiv [Preprint]. 2024 Dec 20:2024.12.17.629035. doi: 10.1101/2024.12.17.629035. bioRxiv. 2024. PMID: 40093150 Free PMC article. Preprint.
-
Single-dose psychedelic enhances cognitive flexibility and reversal learning in mice weeks after administration.Psychedelics (N Y). 2025 Apr 22:10.61373/pp025r.0002. doi: 10.61373/pp025r.0002. Online ahead of print. Psychedelics (N Y). 2025. PMID: 40809535 Free PMC article.
References
-
- Arcelus J, Mitchell AJ, Wales J, Nielsen S. Mortality rates in patients with anorexia nervosa and other eating disorders. A meta-analysis of 36 studies. Arch Gen Psychiatry. 2011;68:724–31. - PubMed
-
- Ferguson CP, La Via MC, Crossan PJ, Kaye WH. Are serotonin selective reuptake inhibitors effective in underweight anorexia nervosa? Int J Eat Disord. 1999;25:11–7. - PubMed
-
- Miles S, Phillipou A, Sumner P, Nedeljkovic M. Cognitive flexibility and the risk of anorexia nervosa: an investigation using self-report and neurocognitive assessments. J Psychiatr Res. 2022;151:531–8. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
