

Higher-order thalamocortical circuits are specified by embryonic cortical progenitor types in the mouse brain
- PMID: 38678557
- PMCID: PMC7617957
- DOI: 10.1016/j.celrep.2024.114157
Higher-order thalamocortical circuits are specified by embryonic cortical progenitor types in the mouse brain
Abstract
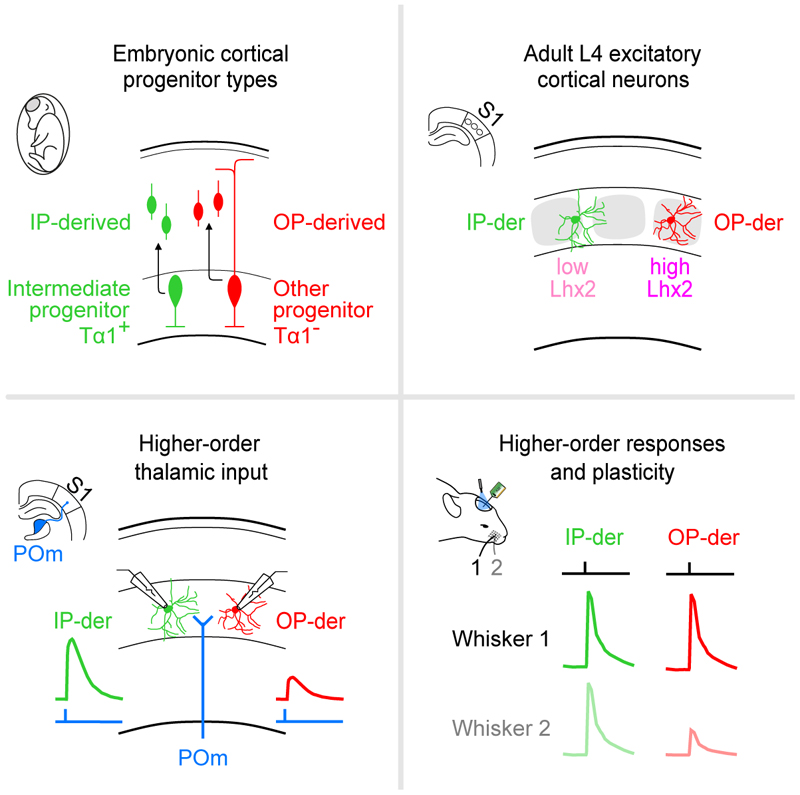
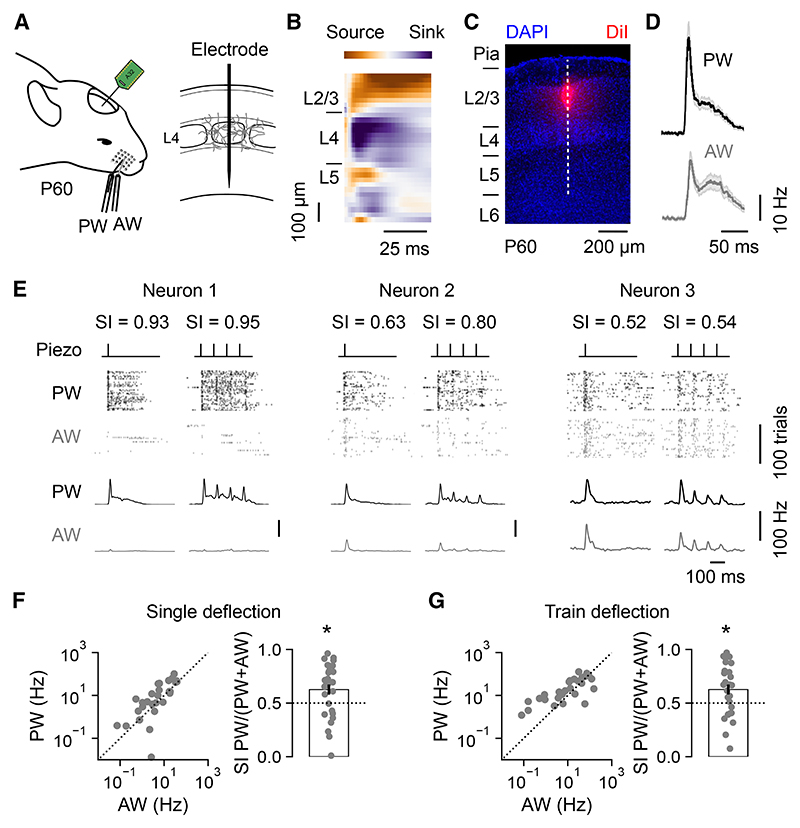
The sensory cortex receives synaptic inputs from both first-order and higher-order thalamic nuclei. First-order inputs relay simple stimulus properties from the periphery, whereas higher-order inputs relay more complex response properties, provide contextual feedback, and modulate plasticity. Here, we reveal that a cortical neuron's higher-order input is determined by the type of progenitor from which it is derived during embryonic development. Within layer 4 (L4) of the mouse primary somatosensory cortex, neurons derived from intermediate progenitors receive stronger higher-order thalamic input and exhibit greater higher-order sensory responses. These effects result from differences in dendritic morphology and levels of the transcription factor Lhx2, which are specified by the L4 neuron's progenitor type. When this mechanism is disrupted, cortical circuits exhibit altered higher-order responses and sensory-evoked plasticity. Therefore, by following distinct trajectories, progenitor types generate diversity in thalamocortical circuitry and may provide a general mechanism for differentially routing information through the cortex.
Keywords: CP: Neuroscience; cortex; development; higher-order; in utero labelling; intermediate progenitor; lineage; optotagging; sensory-evoked plasticity; thalamus.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






Similar articles
-
Peripuberty Is a Sensitive Period for Prefrontal Parvalbumin Interneuron Activity to Impact Adult Cognitive Flexibility.Dev Neurosci. 2025;47(2):127-138. doi: 10.1159/000539584. Epub 2024 Jun 3. Dev Neurosci. 2025. PMID: 38830346 Free PMC article.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
A transcriptional constraint mechanism limits the homeostatic response to activity deprivation in mammalian neocortex.Elife. 2023 Feb 7;12:e74899. doi: 10.7554/eLife.74899. Elife. 2023. PMID: 36749029 Free PMC article.
-
A Reconsideration of the Core and Matrix Classification of Thalamocortical Projections.J Neurosci. 2024 Jun 12;44(24):e0163242024. doi: 10.1523/JNEUROSCI.0163-24.2024. J Neurosci. 2024. PMID: 38866538 Free PMC article. Review.
-
The effect of sample site and collection procedure on identification of SARS-CoV-2 infection.Cochrane Database Syst Rev. 2024 Dec 16;12(12):CD014780. doi: 10.1002/14651858.CD014780. Cochrane Database Syst Rev. 2024. PMID: 39679851 Free PMC article.
Cited by
-
Sensory modality-specific wiring of thalamocortical circuits.Nat Rev Neurosci. 2025 Jul 30. doi: 10.1038/s41583-025-00945-y. Online ahead of print. Nat Rev Neurosci. 2025. PMID: 40745219 Review.
References
-
- Reid RC, Alonso JM. Specificity of monosynaptic connections from thalamus to visual cortex. Nature. 1995;378:281–284. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
