

Identification of Potent and Selective Inhibitors of Acanthamoeba: Structural Insights into Sterol 14α-Demethylase as a Key Drug Target
- PMID: 38683753
- PMCID: PMC11089504
- DOI: 10.1021/acs.jmedchem.4c00303
Identification of Potent and Selective Inhibitors of Acanthamoeba: Structural Insights into Sterol 14α-Demethylase as a Key Drug Target
Abstract
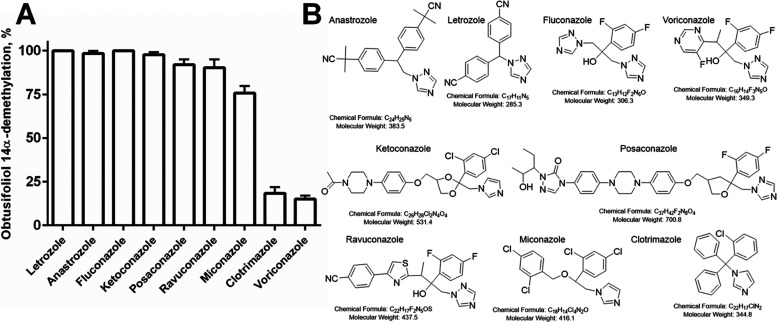
Acanthamoeba are free-living pathogenic protozoa that cause blinding keratitis, disseminated infection, and granulomatous amebic encephalitis, which is generally fatal. The development of efficient and safe drugs is a critical unmet need. Acanthamoeba sterol 14α-demethylase (CYP51) is an essential enzyme of the sterol biosynthetic pathway. Repurposing antifungal azoles for amoebic infections has been reported, but their inhibitory effects on Acanthamoeba CYP51 enzymatic activity have not been studied. Here, we report catalytic properties, inhibition, and structural characterization of CYP51 from Acanthamoeba castellanii. The enzyme displays a 100-fold substrate preference for obtusifoliol over lanosterol, supporting the plant-like cycloartenol-based pathway in the pathogen. The strongest inhibition was observed with voriconazole (1 h IC50 0.45 μM), VT1598 (0.25 μM), and VT1161 (0.20 μM). The crystal structures of A. castellanii CYP51 with bound VT1161 (2.24 Å) and without an inhibitor (1.95 Å), presented here, can be used in the development of azole-based scaffolds to achieve optimal amoebicidal effectiveness.
Conflict of interest statement
The authors declare no competing financial interest.
Figures











Similar articles
-
Characterisation of sterol biosynthesis and validation of 14α-demethylase as a drug target in Acanthamoeba.Sci Rep. 2017 Aug 15;7(1):8247. doi: 10.1038/s41598-017-07495-z. Sci Rep. 2017. PMID: 28811501 Free PMC article.
-
Discovery of a potent, broad-spectrum and in vivo effective deuterated tetrazole CYP51 inhibitor.Eur J Med Chem. 2025 Oct 15;296:117817. doi: 10.1016/j.ejmech.2025.117817. Epub 2025 May 28. Eur J Med Chem. 2025. PMID: 40472486
-
Azole Antifungal Agents To Treat the Human Pathogens Acanthamoeba castellanii and Acanthamoeba polyphaga through Inhibition of Sterol 14α-Demethylase (CYP51).Antimicrob Agents Chemother. 2015 Aug;59(8):4707-13. doi: 10.1128/AAC.00476-15. Epub 2015 May 26. Antimicrob Agents Chemother. 2015. PMID: 26014948 Free PMC article.
-
An overview of azoles targeting sterol 14α-demethylase for antileishmanial therapy.Eur J Med Chem. 2017 Jul 28;135:241-259. doi: 10.1016/j.ejmech.2017.04.044. Epub 2017 Apr 21. Eur J Med Chem. 2017. PMID: 28456033 Review.
-
Management of urinary stones by experts in stone disease (ESD 2025).Arch Ital Urol Androl. 2025 Jun 30;97(2):14085. doi: 10.4081/aiua.2025.14085. Epub 2025 Jun 30. Arch Ital Urol Androl. 2025. PMID: 40583613 Review.
Cited by
-
Unique Structural Features Relate to Evolutionary Adaptation of Cytochrome P450 in the Abyssal Zone.Int J Mol Sci. 2025 Jun 13;26(12):5689. doi: 10.3390/ijms26125689. Int J Mol Sci. 2025. PMID: 40565153 Free PMC article.
-
A First-in-Class Pyrazole-isoxazole Enhanced Antifungal Activity of Voriconazole: Synergy Studies in an Azole-Resistant Candida albicans Strain, Computational Investigation and in Vivo Validation in a Galleria mellonella Fungal Infection Model.J Med Chem. 2024 Aug 22;67(16):14256-14276. doi: 10.1021/acs.jmedchem.4c01109. Epub 2024 Aug 8. J Med Chem. 2024. PMID: 39115219 Free PMC article.
-
Unique structural features in a deep-sea CYP51 relate to high pressure adaptation.Res Sq [Preprint]. 2024 Dec 12:rs.3.rs-5589110. doi: 10.21203/rs.3.rs-5589110/v1. Res Sq. 2024. PMID: 39711526 Free PMC article. Preprint.
-
Identification of Microbial-Based Natural Products as Potential CYP51 Inhibitors for Eumycetoma Treatment: Insights from Molecular Docking, MM-GBSA Calculations, ADMET Analysis, and Molecular Dynamics Simulations.Pharmaceuticals (Basel). 2025 Apr 20;18(4):598. doi: 10.3390/ph18040598. Pharmaceuticals (Basel). 2025. PMID: 40284033 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
