

Developmental fine-tuning of medial superior olive neurons mitigates their predisposition to contralateral sound sources
- PMID: 38683852
- PMCID: PMC11081505
- DOI: 10.1371/journal.pbio.3002586
Developmental fine-tuning of medial superior olive neurons mitigates their predisposition to contralateral sound sources
Abstract
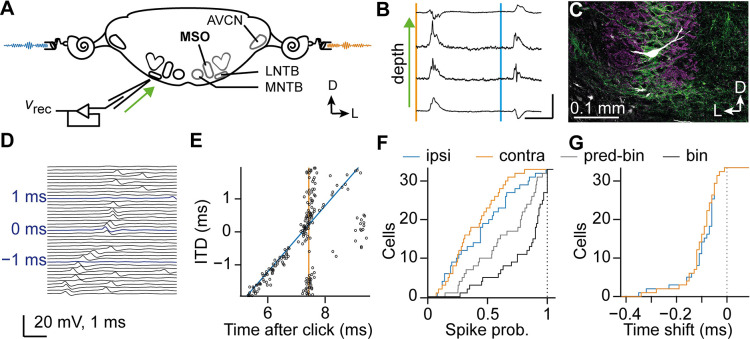
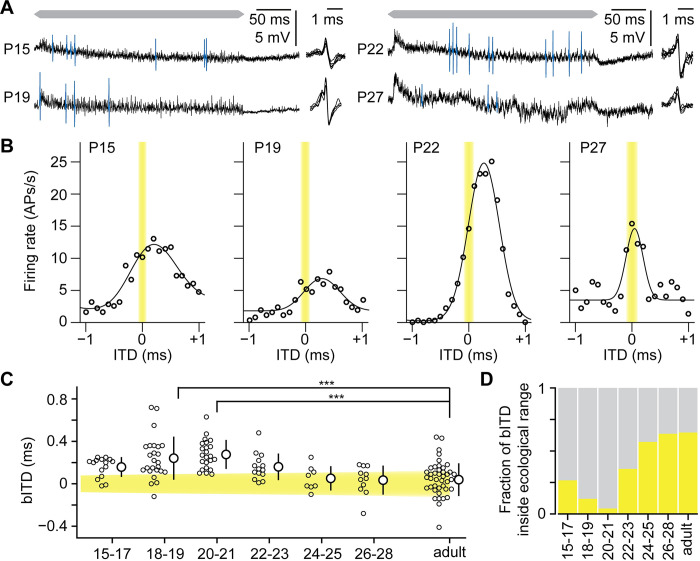
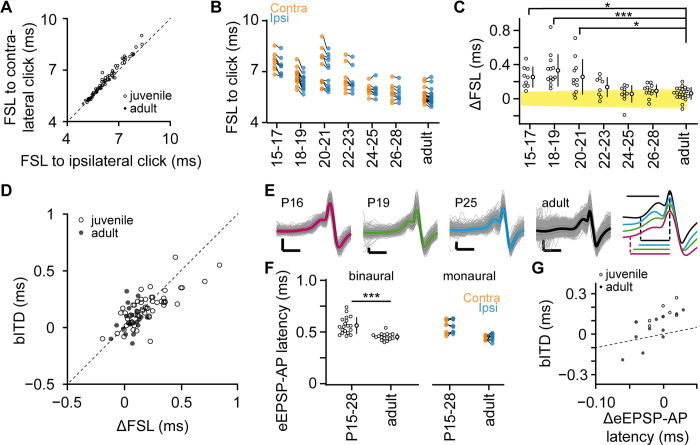
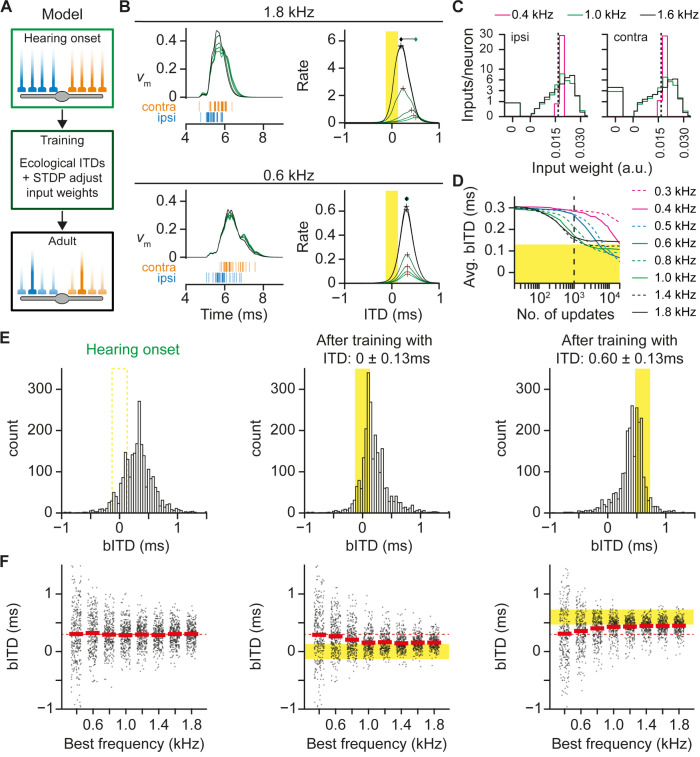
Having two ears enables us to localize sound sources by exploiting interaural time differences (ITDs) in sound arrival. Principal neurons of the medial superior olive (MSO) are sensitive to ITD, and each MSO neuron responds optimally to a best ITD (bITD). In many cells, especially those tuned to low sound frequencies, these bITDs correspond to ITDs for which the contralateral ear leads, and are often larger than the ecologically relevant range, defined by the ratio of the interaural distance and the speed of sound. Using in vivo recordings in gerbils, we found that shortly after hearing onset the bITDs were even more contralaterally leading than found in adult gerbils, and travel latencies for contralateral sound-evoked activity clearly exceeded those for ipsilateral sounds. During the following weeks, both these latencies and their interaural difference decreased. A computational model indicated that spike timing-dependent plasticity can underlie this fine-tuning. Our results suggest that MSO neurons start out with a strong predisposition toward contralateral sounds due to their longer neural travel latencies, but that, especially in high-frequency neurons, this predisposition is subsequently mitigated by differential developmental fine-tuning of the travel latencies.
Copyright: © 2024 Sierksma, Borst. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
