

Discovering a mitochondrion-localized BAHD acyltransferase involved in calystegine biosynthesis and engineering the production of 3β-tigloyloxytropane
- PMID: 38684703
- PMCID: PMC11058270
- DOI: 10.1038/s41467-024-47968-0
Discovering a mitochondrion-localized BAHD acyltransferase involved in calystegine biosynthesis and engineering the production of 3β-tigloyloxytropane
Abstract
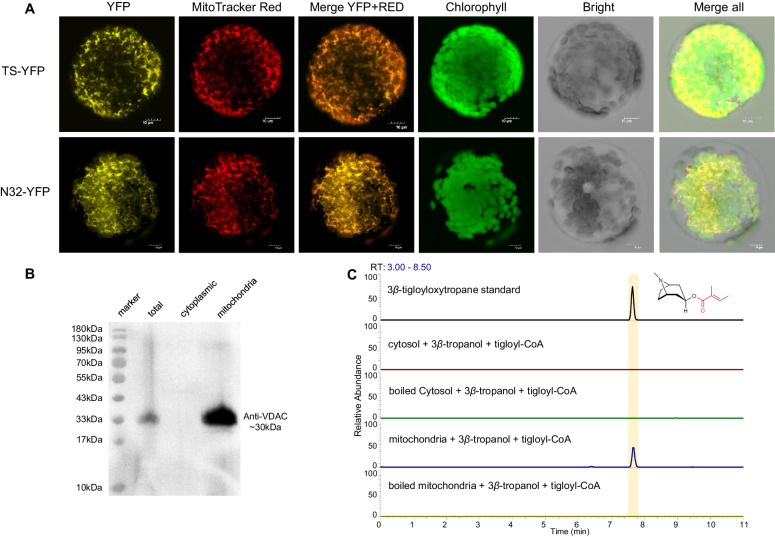
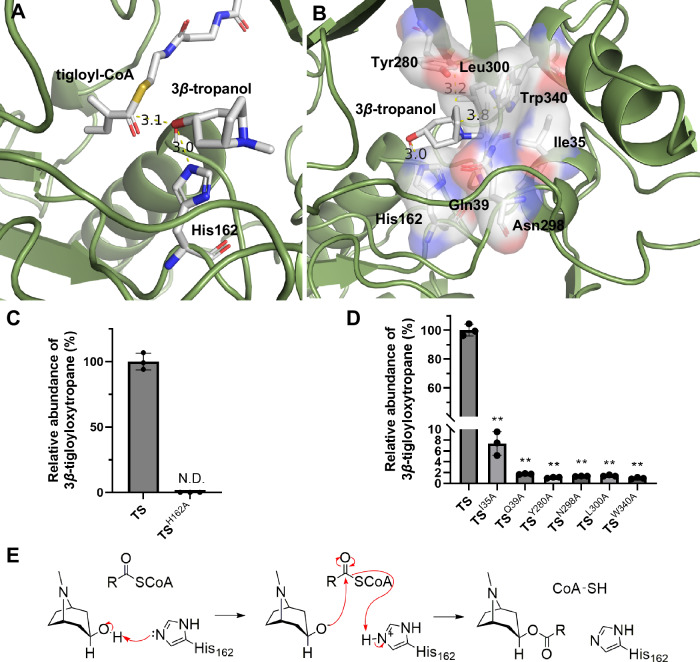
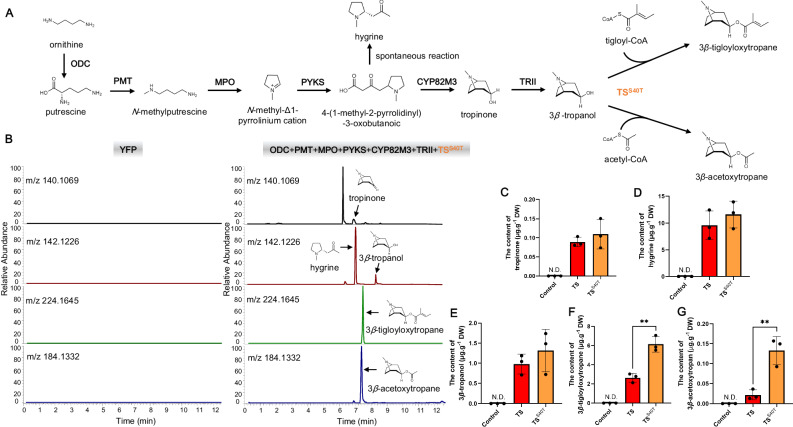
Solanaceous plants produce tropane alkaloids (TAs) via esterification of 3α- and 3β-tropanol. Although littorine synthase is revealed to be responsible for 3α-tropanol esterification that leads to hyoscyamine biosynthesis, the genes associated with 3β-tropanol esterification are unknown. Here, we report that a BAHD acyltransferase from Atropa belladonna, 3β-tigloyloxytropane synthase (TS), catalyzes 3β-tropanol and tigloyl-CoA to form 3β-tigloyloxytropane, the key intermediate in calystegine biosynthesis and a potential drug for treating neurodegenerative disease. Unlike other cytosolic-localized BAHD acyltransferases, TS is localized to mitochondria. The catalytic mechanism of TS is revealed through molecular docking and site-directed mutagenesis. Subsequently, 3β-tigloyloxytropane is synthesized in tobacco. A bacterial CoA ligase (PcICS) is found to synthesize tigloyl-CoA, an acyl donor for 3β-tigloyloxytropane biosynthesis. By expressing TS mutant and PcICS, engineered Escherichia coli synthesizes 3β-tigloyloxytropane from tiglic acid and 3β-tropanol. This study helps to characterize the enzymology and chemodiversity of TAs and provides an approach for producing 3β-tigloyloxytropane.
© 2024. The Author(s).
Conflict of interest statement
On March 1, 2024, J.Z and Z.L., as inventors, applied for patents on the application methods of TS, TSS40T, and PcICS in 3
Figures





Similar articles
-
The BAHD and the bold: the mitochondria's role in alkaloid artistry.Trends Plant Sci. 2024 Dec;29(12):1290-1291. doi: 10.1016/j.tplants.2024.07.012. Epub 2024 Jul 31. Trends Plant Sci. 2024. PMID: 39089907
-
Functional characterization and catalytic activity improvement of BAHD acyltransferase from Celastrus angulatus Maxim.Planta. 2020 Jun 16;252(1):6. doi: 10.1007/s00425-020-03413-2. Planta. 2020. PMID: 32556997
-
A novel bHLH gene responsive to low nitrogen positively regulates the biosynthesis of medicinal tropane alkaloids in Atropa belladonna.Int J Biol Macromol. 2024 May;266(Pt 1):131012. doi: 10.1016/j.ijbiomac.2024.131012. Epub 2024 Mar 23. Int J Biol Macromol. 2024. PMID: 38522709
-
BAHD or SCPL acyltransferase? What a dilemma for acylation in the world of plant phenolic compounds.New Phytol. 2015 Nov;208(3):695-707. doi: 10.1111/nph.13498. Epub 2015 Jun 5. New Phytol. 2015. PMID: 26053460 Review.
-
Tropinone reductase: A comprehensive review on its role as the key enzyme in tropane alkaloids biosynthesis.Int J Biol Macromol. 2023 Dec 31;253(Pt 7):127377. doi: 10.1016/j.ijbiomac.2023.127377. Epub 2023 Oct 14. Int J Biol Macromol. 2023. PMID: 37839598 Review.
Cited by
-
Unraveling the specialized metabolic pathways in medicinal plant genomes: a review.Front Plant Sci. 2024 Dec 24;15:1459533. doi: 10.3389/fpls.2024.1459533. eCollection 2024. Front Plant Sci. 2024. PMID: 39777086 Free PMC article. Review.
-
Engineering scutellarin biosynthesis in Artemisia annua.Plant Cell Rep. 2025 Mar 21;44(4):79. doi: 10.1007/s00299-025-03471-4. Plant Cell Rep. 2025. PMID: 40116969
-
Decoding and reprogramming of the biosynthetic networks of mushroom-derived bioactive type II ganoderic acids in yeast.Cell Discov. 2025 Jul 8;11(1):61. doi: 10.1038/s41421-025-00812-1. Cell Discov. 2025. PMID: 40623979 Free PMC article.
-
Molecular Identification and Functional Characterization of a Putrescine Hydroxycinnamoyltransferase in Suspension Cells of Bamboo (Phyllostachys nigra).Appl Biochem Biotechnol. 2025 Aug 9. doi: 10.1007/s12010-025-05349-3. Online ahead of print. Appl Biochem Biotechnol. 2025. PMID: 40782307
References
-
- Qiu F, et al. Biochemical and metabolic insights into hyoscyamine dehydrogenase. ACS Catal. 2021;11:2912–2924. doi: 10.1021/acscatal.0c04667. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
