

Synergy between Winter Flounder antimicrobial peptides
- PMID: 38686212
- PMCID: PMC11057203
- DOI: 10.1038/s44259-023-00010-7
Synergy between Winter Flounder antimicrobial peptides
Abstract
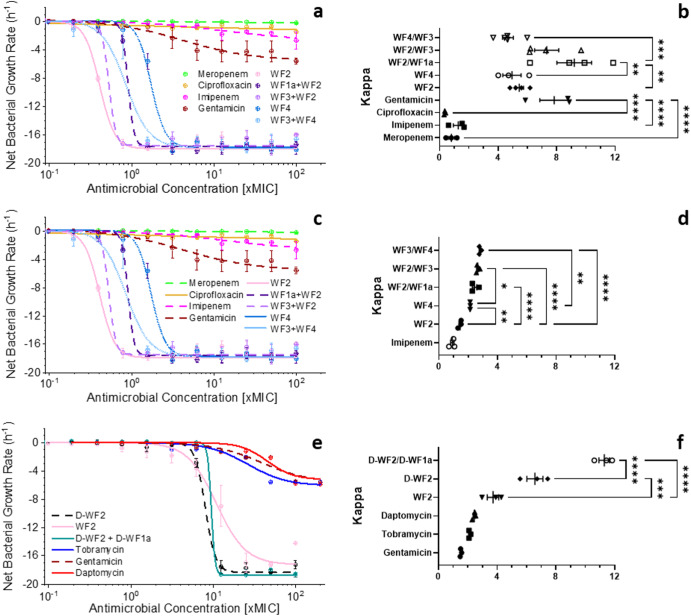
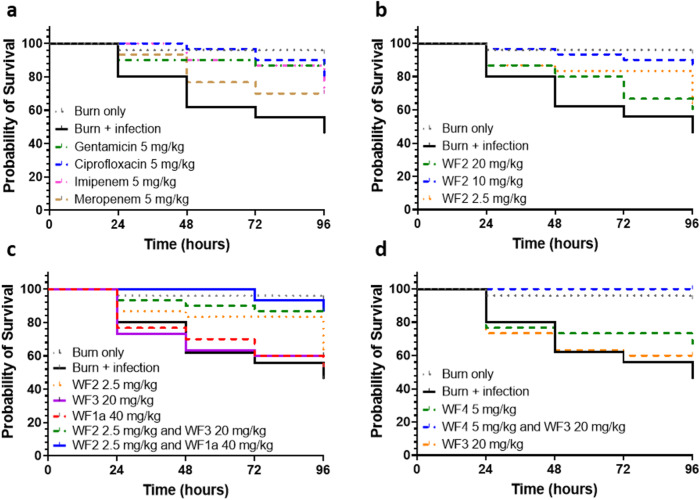
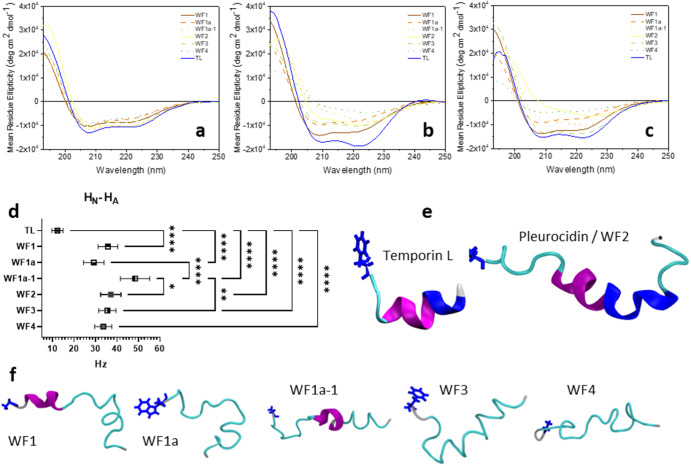
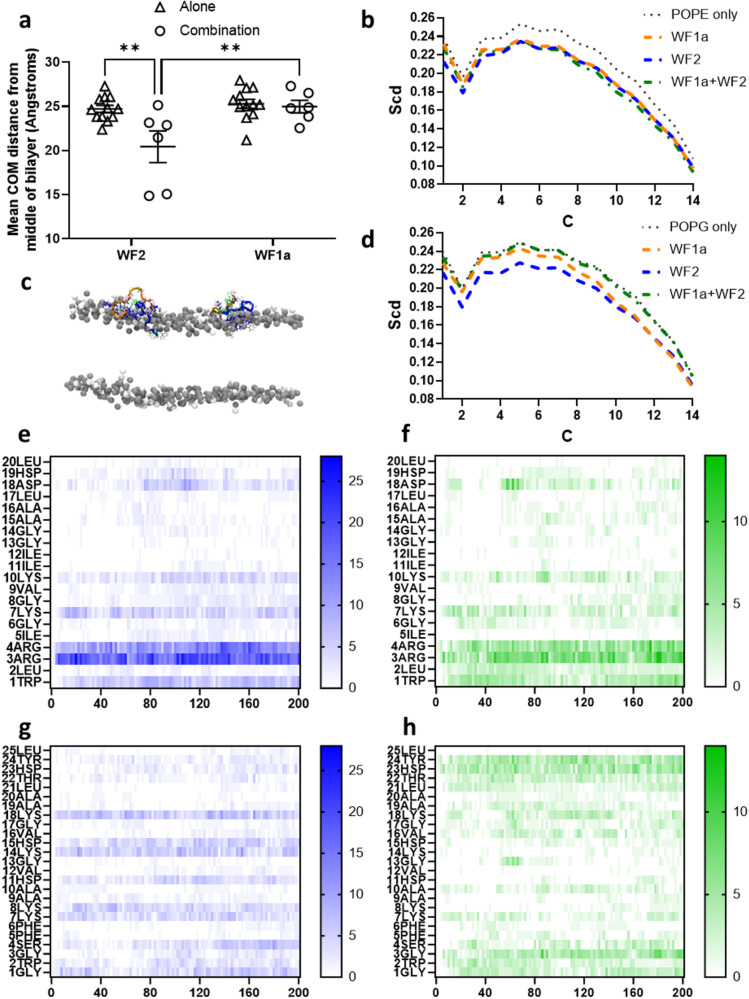
Some antimicrobial peptides (AMPs) have potent bactericidal activity and are being considered as potential alternatives to classical antibiotics. In response to an infection, such AMPs are often produced in animals alongside other peptides with low or no perceivable antimicrobial activity, whose role is unclear. Here we show that six AMPs from the Winter Flounder (WF) act in synergy against a range of bacterial pathogens and provide mechanistic insights into how this increases the cooperativity of the dose-dependent bactericidal activity and potency that enable therapy. Only two WF AMPs have potent antimicrobial activity when used alone but we find a series of two-way combinations, involving peptides which otherwise have low or no activity, yield potent antimicrobial activity. Weakly active WF AMPs modulate the membrane interactions of the more potent WF AMPs and enable therapy in a model of Acinetobacter baumannii burn wound infection. The observed synergy and emergent behaviour may explain the evolutionary benefits of producing a family of related peptides and are attractive properties to consider when developing AMPs towards clinical applications.
Keywords: Antibiotics; Computational models; Peptides; Pharmacodynamics.
© The Author(s) 2023.
Conflict of interest statement
Competing interestsThe authors declare no competing interests.
Figures


References
-
- Jangir, P. K., Ogunlana, L. & MacLean, R. C. Evolutionary constraints on the acquisition of antimicrobial peptide resistance in bacterial pathogens. Trends Microbiol.29, 1058–1061 (2021). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
