

Innate immune activation restricts priming and protective efficacy of the radiation-attenuated PfSPZ malaria vaccine
- PMID: 38687615
- PMCID: PMC11382880
- DOI: 10.1172/jci.insight.167408
Innate immune activation restricts priming and protective efficacy of the radiation-attenuated PfSPZ malaria vaccine
Abstract
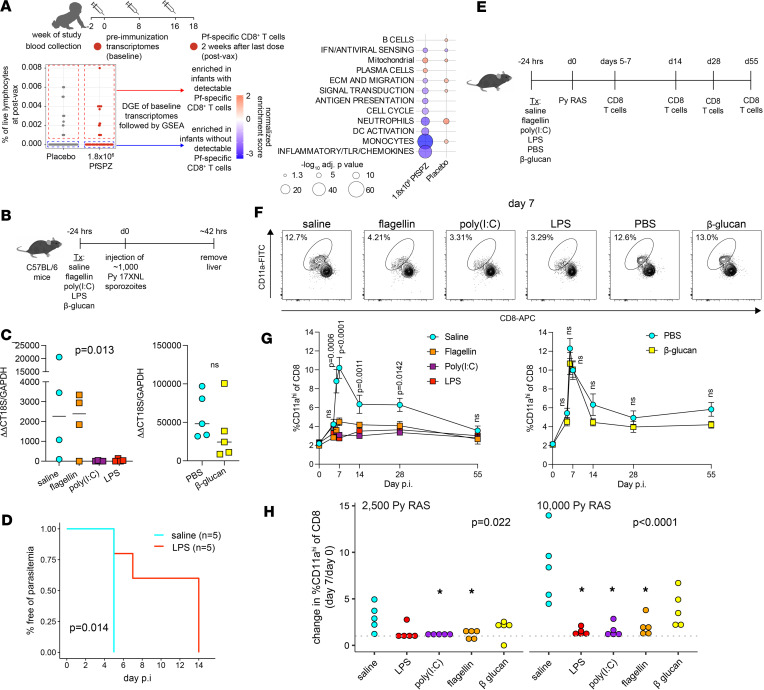
A systems analysis was conducted to determine the potential molecular mechanisms underlying differential immunogenicity and protective efficacy results of a clinical trial of the radiation-attenuated whole-sporozoite PfSPZ vaccine in African infants. Innate immune activation and myeloid signatures at prevaccination baseline correlated with protection from P. falciparum parasitemia in placebo controls. These same signatures were associated with susceptibility to parasitemia among infants who received the highest and most protective PfSPZ vaccine dose. Machine learning identified spliceosome, proteosome, and resting DC signatures as prevaccination features predictive of protection after highest-dose PfSPZ vaccination, whereas baseline circumsporozoite protein-specific (CSP-specific) IgG predicted nonprotection. Prevaccination innate inflammatory and myeloid signatures were associated with higher sporozoite-specific IgG Ab response but undetectable PfSPZ-specific CD8+ T cell responses after vaccination. Consistent with these human data, innate stimulation in vivo conferred protection against infection by sporozoite injection in malaria-naive mice while diminishing the CD8+ T cell response to radiation-attenuated sporozoites. These data suggest a dichotomous role of innate stimulation for malaria protection and induction of protective immunity by whole-sporozoite malaria vaccines. The uncoupling of vaccine-induced protective immunity achieved by Abs from more protective CD8+ T cell responses suggests that PfSPZ vaccine efficacy in malaria-endemic settings may be constrained by opposing antigen presentation pathways.
Keywords: Adaptive immunity; Infectious disease; Innate immunity; Malaria; Vaccines.
Figures








References
-
- RTS S Clinical Trials Partnership. Efficacy and safety of RTS,S/AS01 malaria vaccine with or without a booster dose in infants and children in Africa: final results of a phase 3, individually randomised, controlled trial. Lancet. 2015;386(9988):31–45. doi: 10.1016/S0140-6736(15)60721-8. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
