

Sleep pressure modulates single-neuron synapse number in zebrafish
- PMID: 38693264
- PMCID: PMC11096099
- DOI: 10.1038/s41586-024-07367-3
Sleep pressure modulates single-neuron synapse number in zebrafish
Abstract
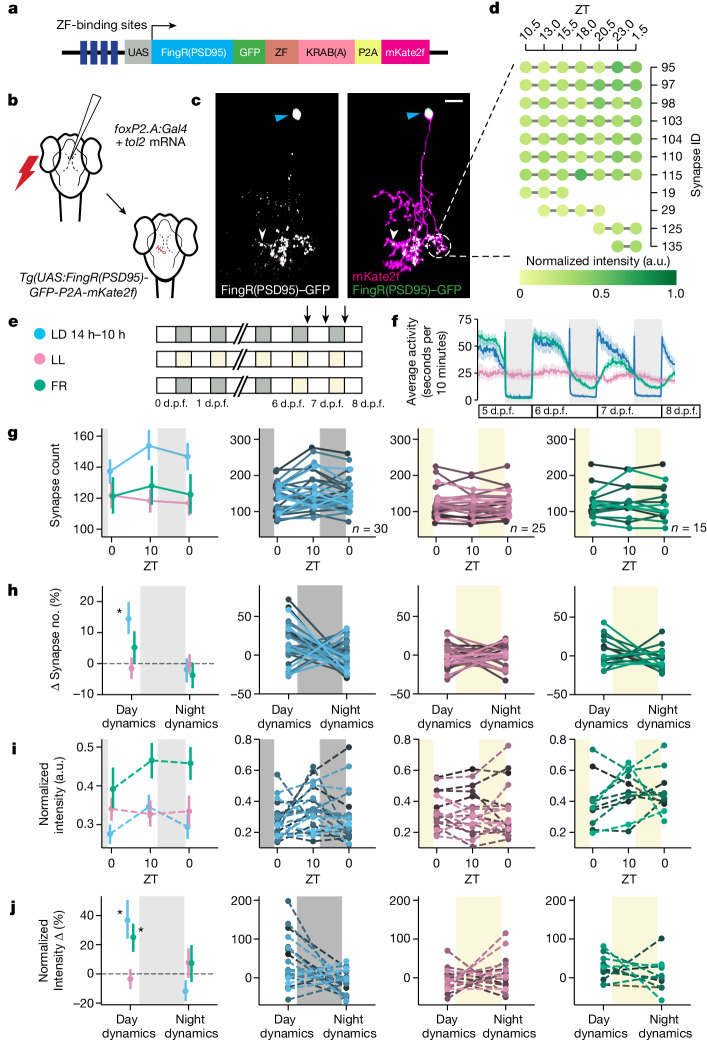
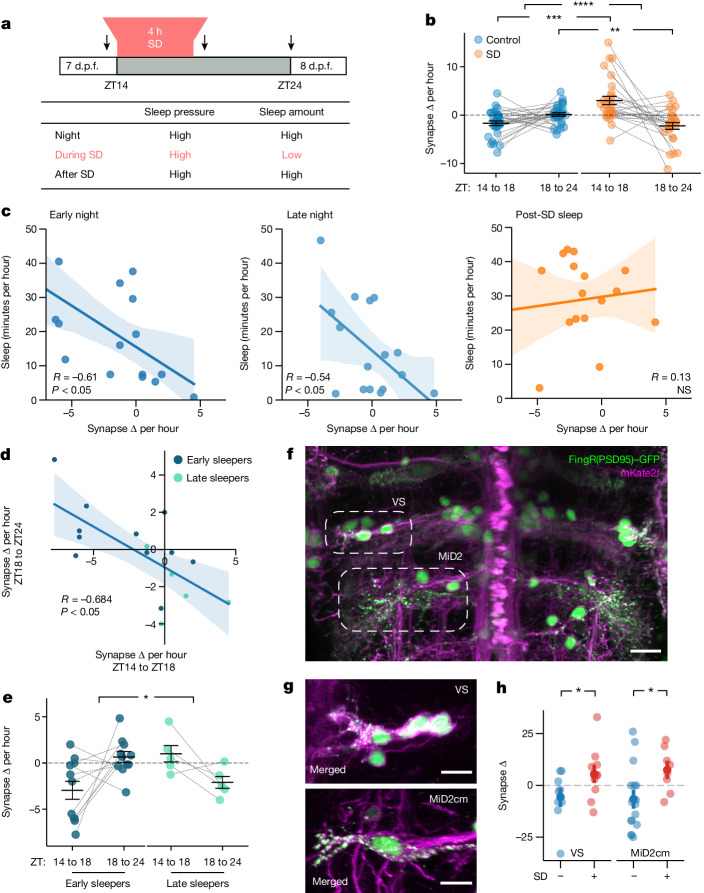
Sleep is a nearly universal behaviour with unclear functions1. The synaptic homeostasis hypothesis proposes that sleep is required to renormalize the increases in synaptic number and strength that occur during wakefulness2. Some studies examining either large neuronal populations3 or small patches of dendrites4 have found evidence consistent with the synaptic homeostasis hypothesis, but whether sleep merely functions as a permissive state or actively promotes synaptic downregulation at the scale of whole neurons is unclear. Here, by repeatedly imaging all excitatory synapses on single neurons across sleep-wake states of zebrafish larvae, we show that synapses are gained during periods of wake (either spontaneous or forced) and lost during sleep in a neuron-subtype-dependent manner. However, synapse loss is greatest during sleep associated with high sleep pressure after prolonged wakefulness, and lowest in the latter half of an undisrupted night. Conversely, sleep induced pharmacologically during periods of low sleep pressure is insufficient to trigger synapse loss unless adenosine levels are boosted while noradrenergic tone is inhibited. We conclude that sleep-dependent synapse loss is regulated by sleep pressure at the level of the single neuron and that not all sleep periods are equally capable of fulfilling the functions of synaptic homeostasis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures













References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
