

The type of inhibition provided by thalamic interneurons alters the input selectivity of thalamocortical neurons
- PMID: 38694514
- PMCID: PMC11061260
- DOI: 10.1016/j.crneur.2024.100130
The type of inhibition provided by thalamic interneurons alters the input selectivity of thalamocortical neurons
Abstract
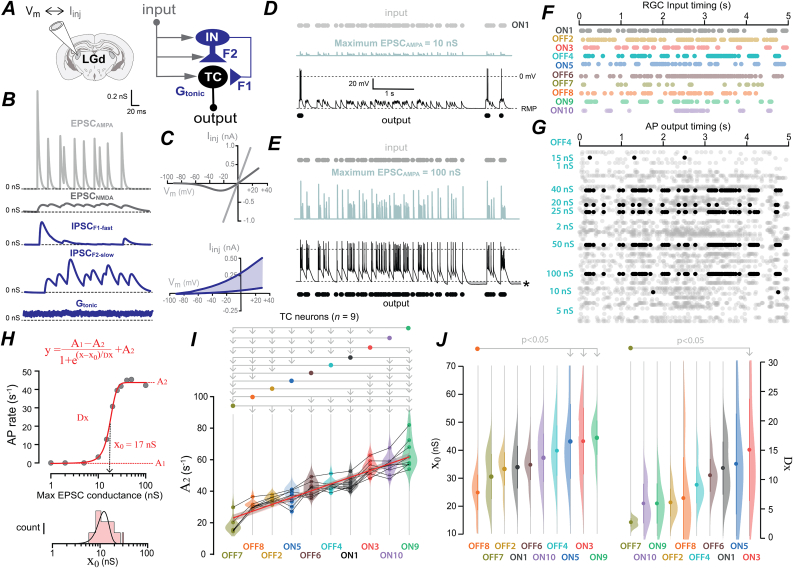
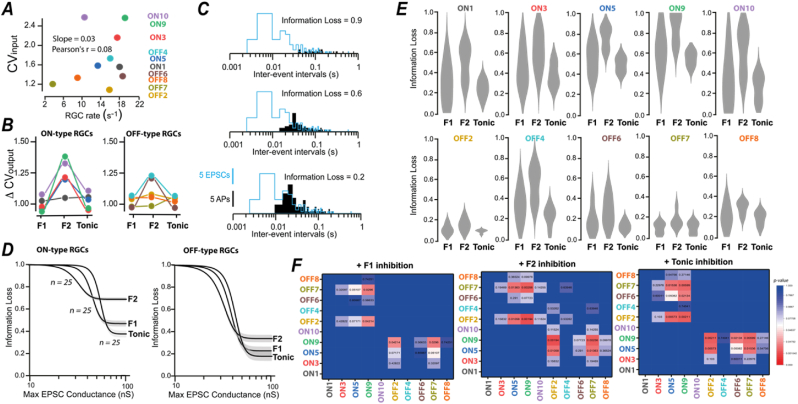
A fundamental problem in neuroscience is how neurons select for their many inputs. A common assumption is that a neuron's selectivity is largely explained by differences in excitatory synaptic input weightings. Here we describe another solution to this important problem. We show that within the first order visual thalamus, the type of inhibition provided by thalamic interneurons has the potential to alter the input selectivity of thalamocortical neurons. To do this, we developed conductance injection protocols to compare how different types of synchronous and asynchronous GABA release influence thalamocortical excitability in response to realistic patterns of retinal ganglion cell input. We show that the asynchronous GABA release associated with tonic inhibition is particularly efficient at maintaining information content, ensuring that thalamocortical neurons can distinguish between their inputs. We propose a model where alterations in GABA release properties results in rapid changes in input selectivity without requiring structural changes in the network.
© 2024 The Authors.
Conflict of interest statement
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests. This research was funded by the Biotechnology and Biological Sciences Research Council (BB/R007659/1)
Figures





References
-
- Acuna-Goycolea C., Brenowitz S.D., Regehr W.G. Active dendritic conductances dynamically regulate GABA release from thalamic Interneurons. Neuron. 2008;57:420–431. - PubMed
-
- Arcelli P., Frassoni C., Regondi M.C., De Biasi S., Spreafico R. GABAergic neurons in mammalian thalamus: a marker of thalamic complexity? Brain Res. Bull. 1997;42:27–37. - PubMed
-
- Bakken T.E., van Velthoven C.T., Menon V., Hodge R.D., Yao Z., Nguyen T.N., Graybuck L.T., Horwitz G.D., Bertagnolli D., Goldy J., Yanny A.M., Garren E., Parry S., Casper T., Shehata S.I., Barkan E.R., Szafer A., Levi B.P., Dee N., Smith K.A., Sunkin S.M., Bernard A., Phillips J., Hawrylycz M.J., Koch C., Murphy G.J., Lein E., Zeng H., Tasic B. Single-cell and single-nucleus RNA-seq uncovers shared and distinct axes of variation in dorsal LGN neurons in mice, non-human primates, and humans. Elife. 2021;10 - PMC - PubMed
-
- Barbaresi P., Spreafico R., Frassoni C., Rustioni A. GABAergic neurons are present in the dorsal column nuclei but not in the ventroposterior complex of rats. Brain Res. 1986;382:305–326. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
