

A novel reporter for helicase activity in translation uncovers DDX3X interactions
- PMID: 38697667
- PMCID: PMC11251518
- DOI: 10.1261/rna.079837.123
A novel reporter for helicase activity in translation uncovers DDX3X interactions
Abstract
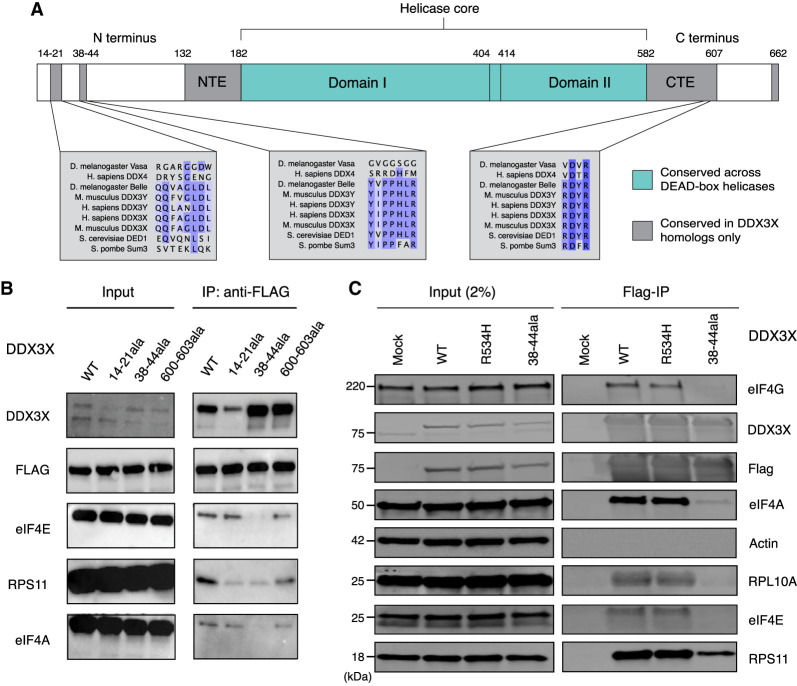
DDX3X regulates the translation of a subset of human transcripts containing complex 5' untranslated regions (5' UTRs). In this study, we developed the helicase activity reporter for translation (HART), which uses DDX3X-sensitive 5' UTRs to measure DDX3X-mediated translational activity in cells. To directly measure RNA structure in DDX3X-dependent mRNAs, we used SHAPE-MaP to determine the secondary structures present in DDX3X-sensitive 5' UTRs and then used HART to investigate how sequence alterations influence DDX3X sensitivity. Additionally, we identified residues 38-44 as potential mediators of DDX3X's interaction with the translational machinery. HART revealed that both DDX3X's association with the translational machinery and its helicase activity are required for its function in promoting the translation of DDX3X-sensitive 5' UTRs. These findings suggest DDX3X plays a crucial role in regulating translation through its interaction with the translational machinery during ribosome scanning and establish the HART reporter as a robust, lentivirally encoded, colorimetric measurement of DDX3X-dependent translation in cells.
Keywords: RNA helicases; RNA structure; reporter genes; translational control.
© 2024 Wilkins et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures





Update of
-
Determinants of DDX3X sensitivity uncovered using a helicase activity in translation reporter.bioRxiv [Preprint]. 2023 Sep 14:2023.09.14.557805. doi: 10.1101/2023.09.14.557805. bioRxiv. 2023. Update in: RNA. 2024 Jul 16;30(8):1041-1057. doi: 10.1261/rna.079837.123. PMID: 37745530 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials