

Assessment of the hypothetical protein BB0616 in the murine infection of Borrelia burgdorferi
- PMID: 38700336
- PMCID: PMC11237664
- DOI: 10.1128/iai.00090-24
Assessment of the hypothetical protein BB0616 in the murine infection of Borrelia burgdorferi
Abstract
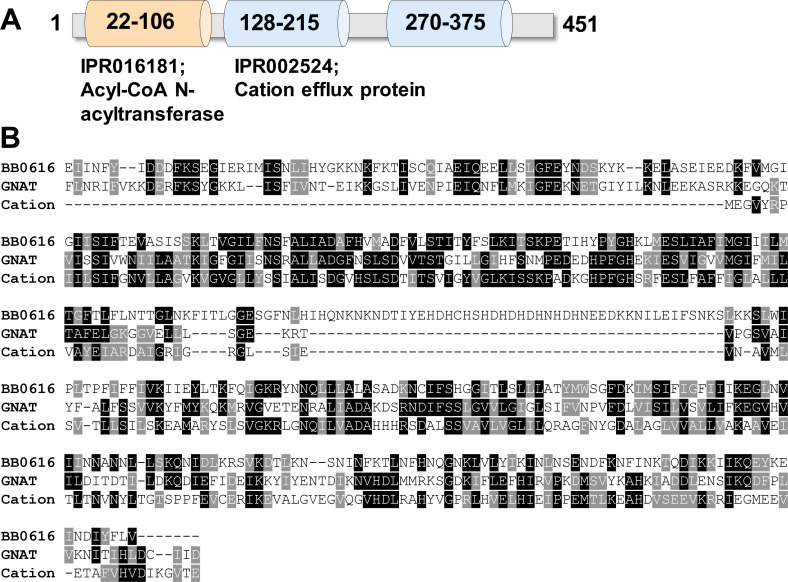
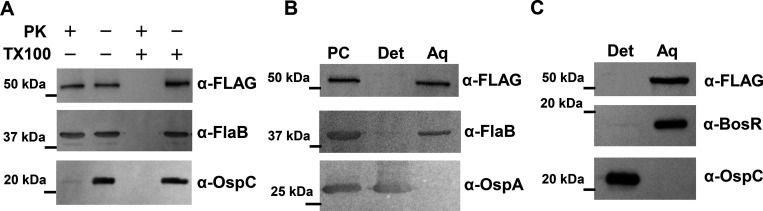
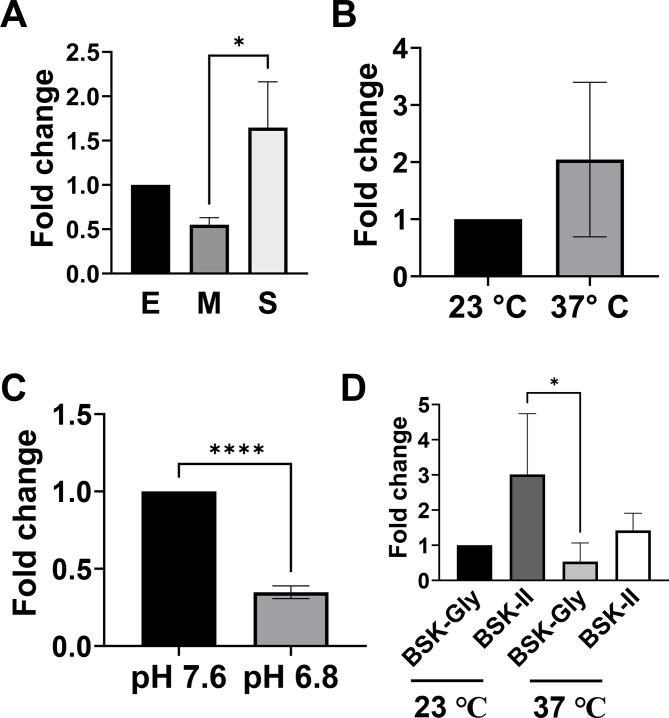
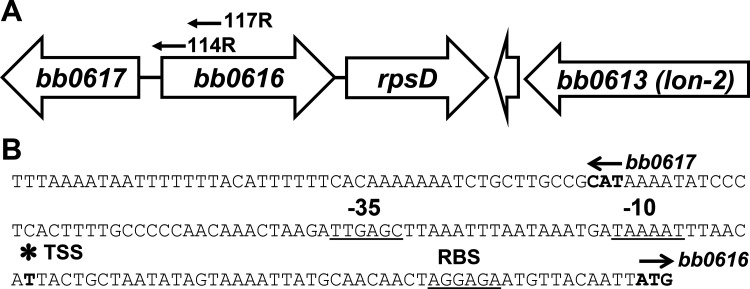
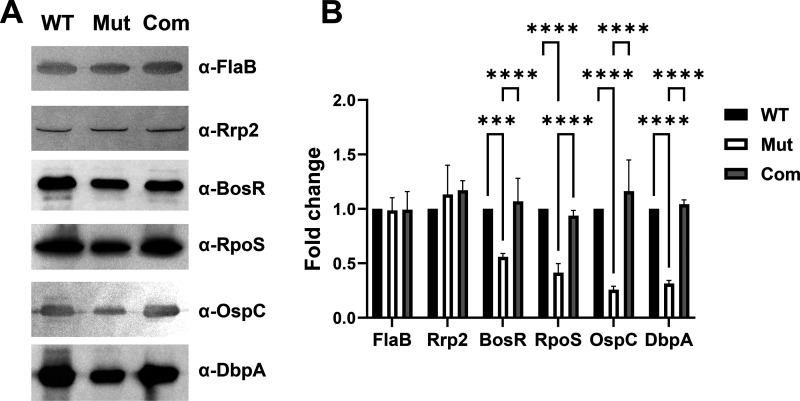
bb0616 of Borrelia burgdorferi, the Lyme disease pathogen, encodes a hypothetical protein of unknown function. In this study, we showed that BB0616 was not surface-exposed or associated with the membrane through localization analyses using proteinase K digestion and cell partitioning assays. The expression of bb0616 was influenced by a reduced pH but not by growth phases, elevated temperatures, or carbon sources during in vitro cultivation. A transcriptional start site for bb0616 was identified by using 5' rapid amplification of cDNA ends, which led to the identification of a functional promoter in the 5' regulatory region upstream of bb0616. By analyzing a bb0616-deficient mutant and its isogenic complemented counterparts, we found that the infectivity potential of the mutant was significantly attenuated. The inactivation of bb0616 displayed no effect on borrelial growth in the medium or resistance to oxidative stress, but the mutant was significantly more susceptible to osmotic stress. In addition, the production of global virulence regulators such as BosR and RpoS as well as virulence-associated outer surface lipoproteins OspC and DbpA was reduced in the mutant. These phenotypes were fully restored when gene mutation was complemented with a wild-type copy of bb0616. Based on these findings, we concluded that the hypothetical protein BB0616 is required for the optimal infectivity of B. burgdorferi, potentially by impacting B. burgdorferi virulence gene expression as well as survival of the spirochete under stressful conditions.
Keywords: Borrelia burgdorferi; Lyme disease; RpoS; pathogenesis; regulation of gene expression; virulence regulation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
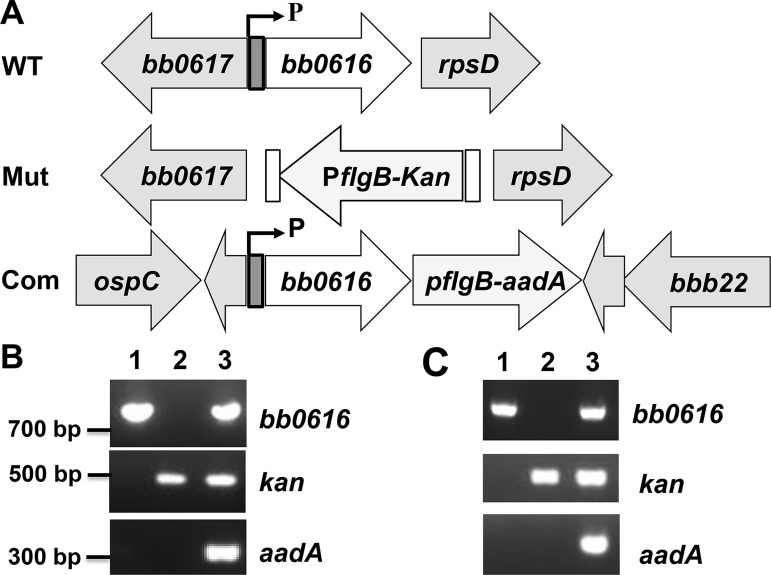
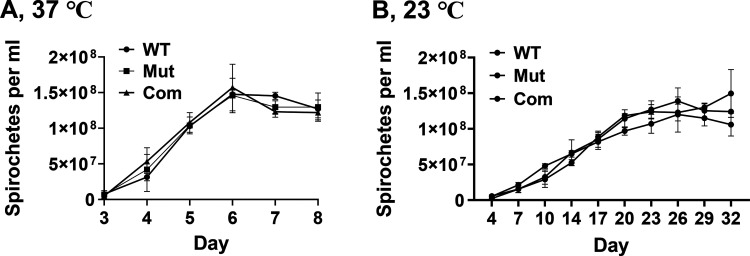
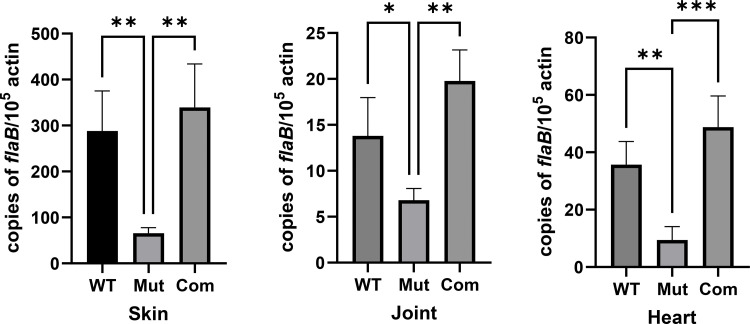
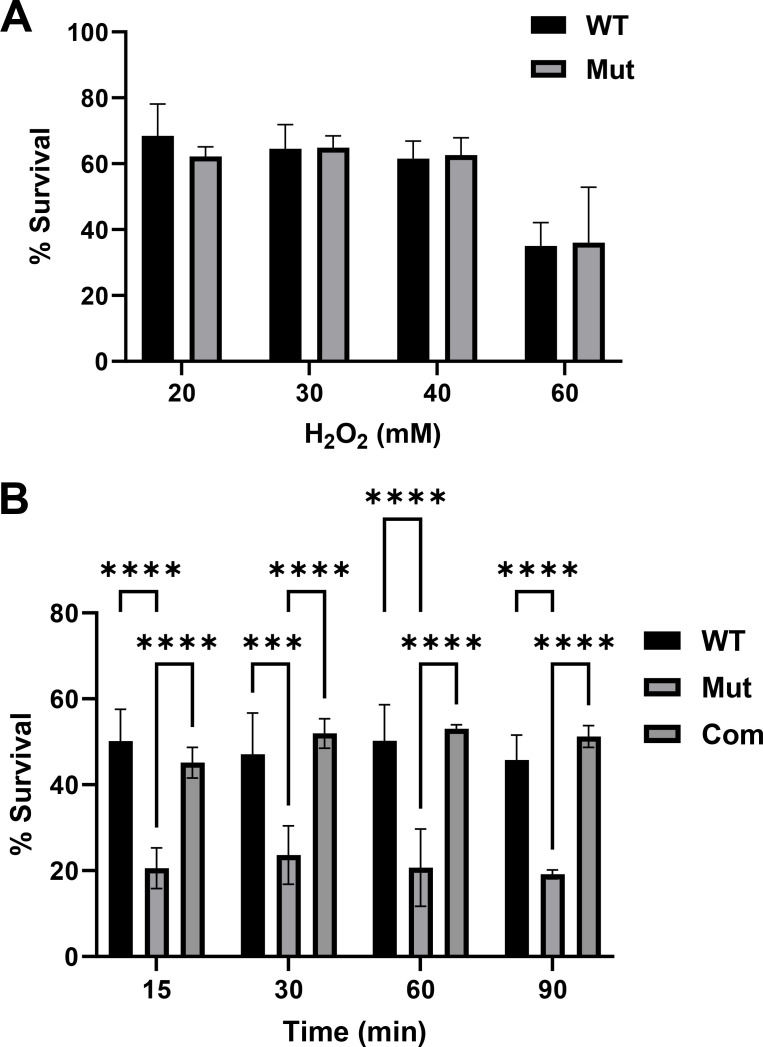
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
