

Intermittent glucocorticoid treatment improves muscle metabolism via the PGC1α/Lipin1 axis in an aging-related sarcopenia model
- PMID: 38702076
- PMCID: PMC11142738
- DOI: 10.1172/JCI177427
Intermittent glucocorticoid treatment improves muscle metabolism via the PGC1α/Lipin1 axis in an aging-related sarcopenia model
Abstract
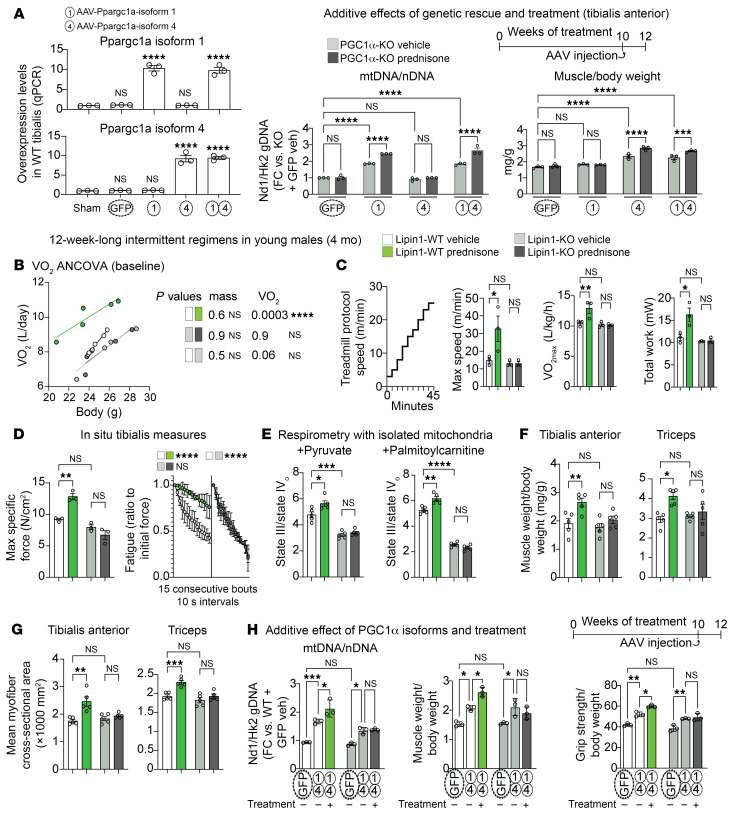
Sarcopenia burdens the older population through loss of muscle energy and mass, yet treatments to functionally rescue both parameters are lacking. The glucocorticoid prednisone remodels muscle metabolism on the basis of frequency of intake, but its mechanisms in sarcopenia are unknown. We found that once-weekly intermittent prednisone administration rescued muscle quality in aged 24-month-old mice to a level comparable to that seen in young 4-month-old mice. We discovered an age- and sex-independent glucocorticoid receptor transactivation program in muscle encompassing peroxisome proliferator-activated receptor γ coactivator 1 α (PGC1α) and its cofactor Lipin1. Treatment coordinately improved mitochondrial abundance through isoform 1 and muscle mass through isoform 4 of the myocyte-specific PGC1α, which was required for the treatment-driven increase in carbon shuttling from glucose oxidation to amino acid biogenesis. We also probed myocyte-specific Lipin1 as a nonredundant factor coaxing PGC1α upregulation to the stimulation of both oxidative and anabolic effects. Our study unveils an aging-resistant druggable program in myocytes for the coordinated rescue of energy and mass in sarcopenia.
Keywords: Aging; Epigenetics; Mitochondria; Muscle; Muscle biology.
Conflict of interest statement
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
