

Structures, mechanisms and applications of RNA-centric CRISPR-Cas13
- PMID: 38702571
- PMCID: PMC11375968
- DOI: 10.1038/s41589-024-01593-6
Structures, mechanisms and applications of RNA-centric CRISPR-Cas13
Abstract
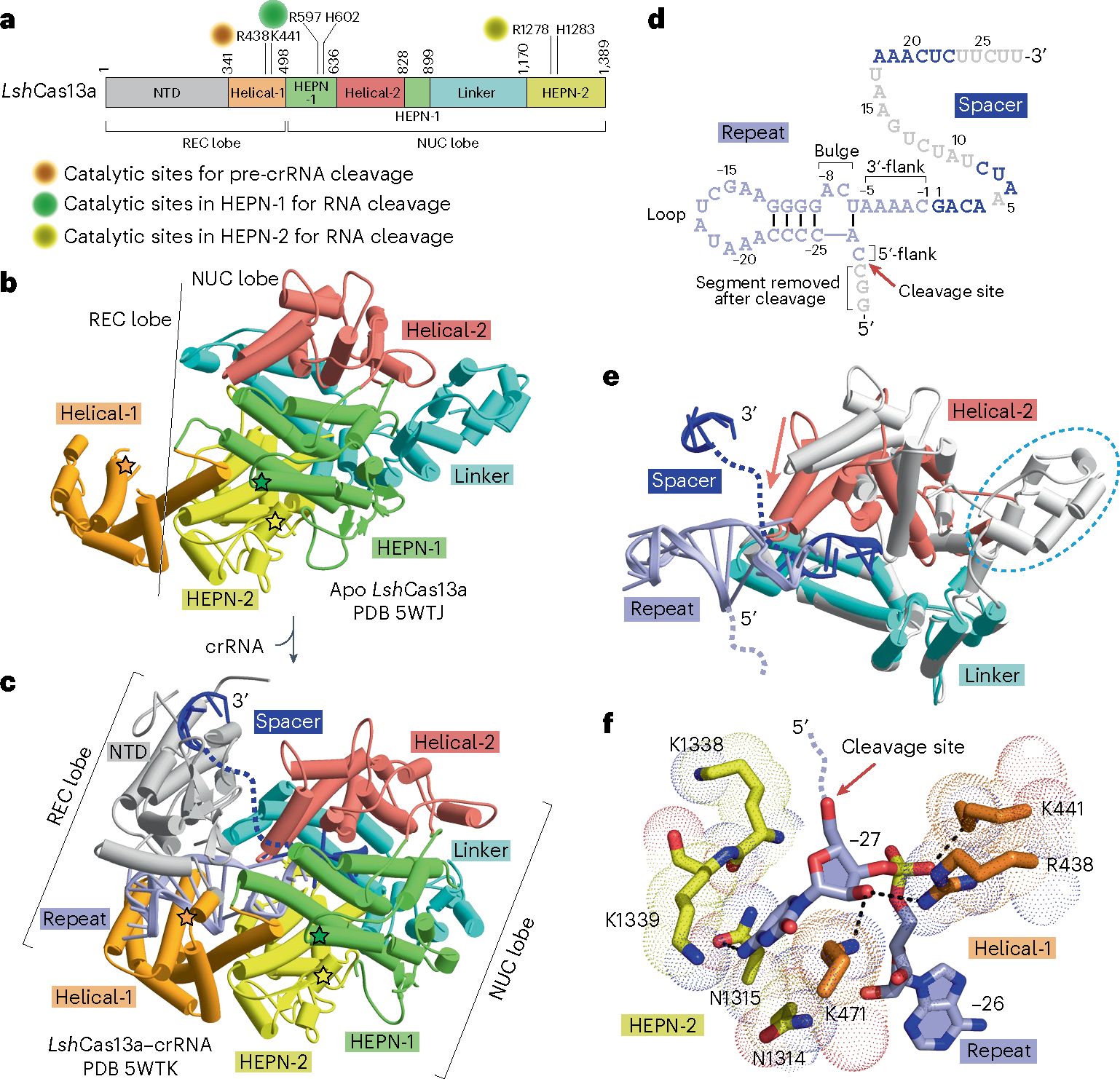
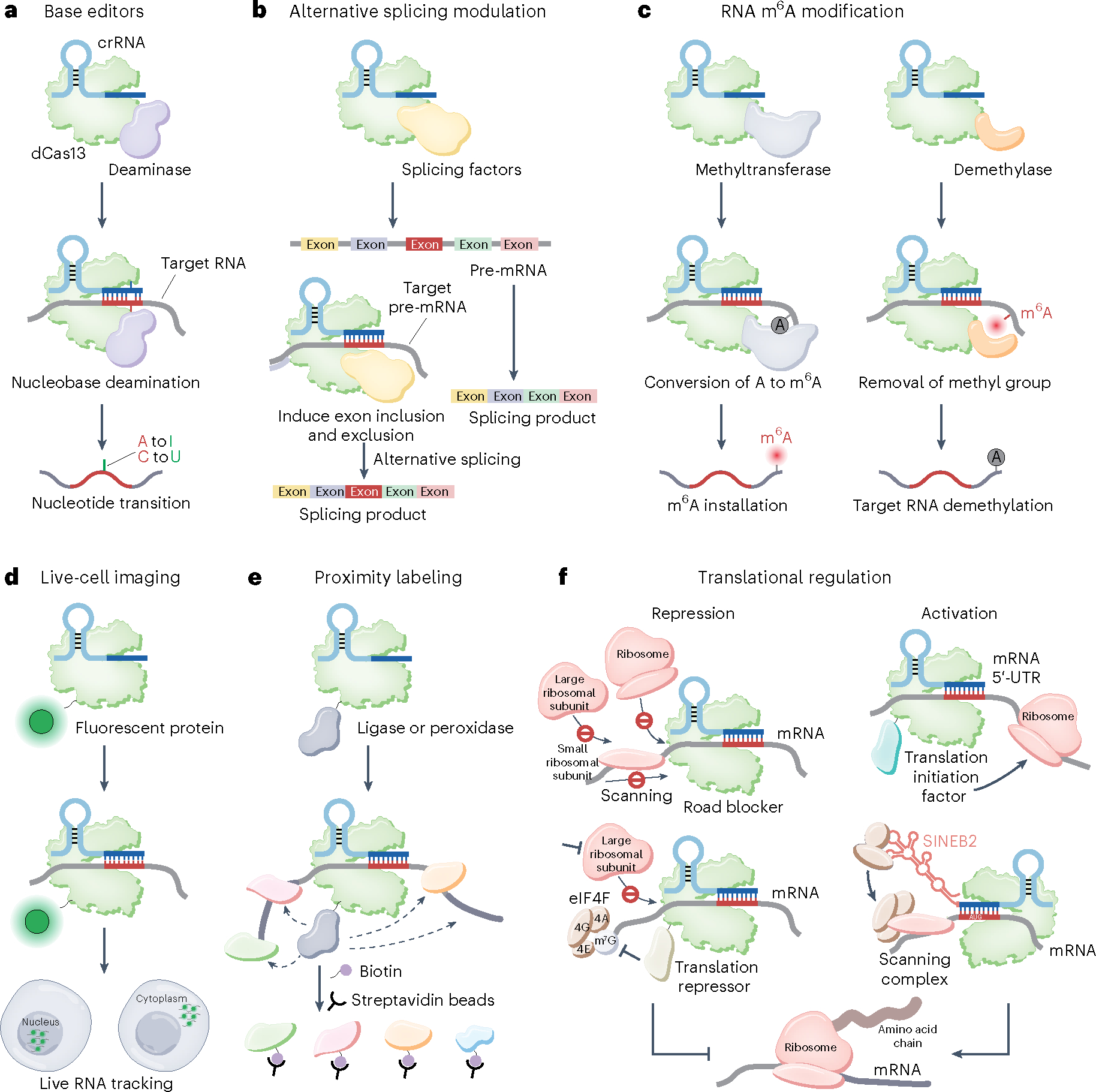
Prokaryotes are equipped with a variety of resistance strategies to survive frequent viral attacks or invading mobile genetic elements. Among these, CRISPR-Cas surveillance systems are abundant and have been studied extensively. This Review focuses on CRISPR-Cas type VI Cas13 systems that use single-subunit RNA-guided Cas endonucleases for targeting and subsequent degradation of foreign RNA, thereby providing adaptive immunity. Notably, distinct from single-subunit DNA-cleaving Cas9 and Cas12 systems, Cas13 exhibits target RNA-activated substrate RNase activity. This Review outlines structural, biochemical and cell biological studies toward elucidation of the unique structural and mechanistic principles underlying surveillance effector complex formation, precursor CRISPR RNA (pre-crRNA) processing, self-discrimination and RNA degradation in Cas13 systems as well as insights into suppression by bacteriophage-encoded anti-CRISPR proteins and regulation by endogenous accessory proteins. Owing to its programmable ability for RNA recognition and cleavage, Cas13 provides powerful RNA targeting, editing, detection and imaging platforms with emerging biotechnological and therapeutic applications.
© 2024. Springer Nature America, Inc.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
- AI141507/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01 GM129430/GM/NIGMS NIH HHS/United States
- GM129430/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- P30 CA008748/CA/NCI NIH HHS/United States
- R01 AI141507/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
