

Interactions between chaperone and energy storage networks during the evolution of Legionella pneumophila under heat shock
- PMID: 38708341
- PMCID: PMC11067923
- DOI: 10.7717/peerj.17197
Interactions between chaperone and energy storage networks during the evolution of Legionella pneumophila under heat shock
Abstract
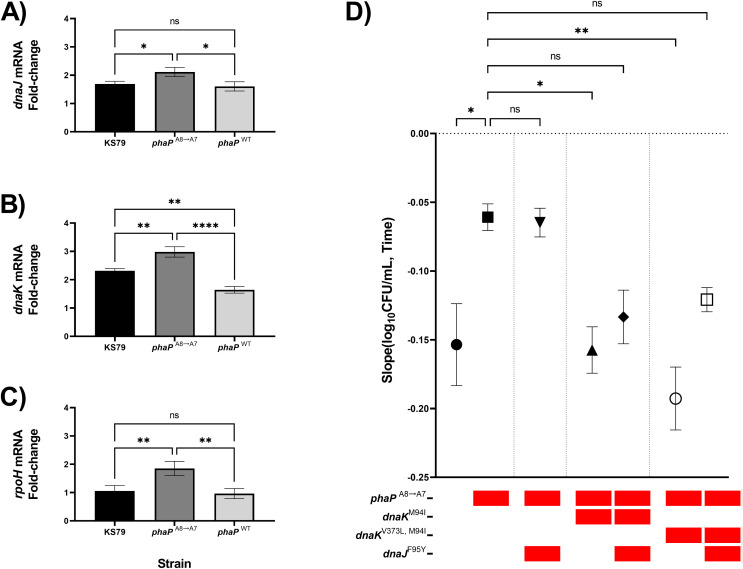
Waterborne transmission of the bacterium Legionella pneumophila has emerged as a major cause of severe nosocomial infections of major public health impact. The major route of transmission involves the uptake of aerosolized bacteria, often from the contaminated hot water systems of large buildings. Public health regulations aimed at controlling the mesophilic pathogen are generally concerned with acute pasteurization and maintaining high temperatures at the heating systems and throughout the plumbing of hot water systems, but L. pneumophila is often able to survive these treatments due to both bacterium-intrinsic and environmental factors. Previous work has established an experimental evolution system to model the observations of increased heat resistance in repeatedly but unsuccessfully pasteurized L. pneumophila populations. Here, we show rapid fixation of novel alleles in lineages selected for resistance to heat shock and shifts in mutational profile related to increases in the temperature of selection. Gene-level and nucleotide-level parallelisms between independently-evolving lineages show the centrality of the DnaJ/DnaK chaperone system in the heat resistance of L. pneumophila. Inference of epistatic interactions through reverse genetics shows an unexpected interaction between DnaJ/DnaK and the polyhydroxybutyrate-accumulation energy storage mechanism used by the species to survive long-term starvation in low-nutrient environments.
Keywords: Experimental evolution; Heat shock proteins; Heat shock response; Legionella pneumophila; Pasteurization; Phasin protein, bacteria; Poly(hydroxybutyrates).
© 2024 Liang and Faucher.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



Similar articles
-
Development of heat-shock resistance in Legionella pneumophila modeled by experimental evolution.Appl Environ Microbiol. 2023 Sep 28;89(9):e0066623. doi: 10.1128/aem.00666-23. Epub 2023 Sep 5. Appl Environ Microbiol. 2023. PMID: 37668382 Free PMC article.
-
Effect of heat shock on hot water plumbing microbiota and Legionella pneumophila control.Microbiome. 2018 Feb 9;6(1):30. doi: 10.1186/s40168-018-0406-7. Microbiome. 2018. PMID: 29426363 Free PMC article.
-
Controlling Legionella pneumophila growth in hot water systems by reducing dissolved oxygen levels.J Occup Environ Hyg. 2024 Mar-Apr;21(4):259-269. doi: 10.1080/15459624.2024.2313580. Epub 2024 Mar 6. J Occup Environ Hyg. 2024. PMID: 38447033
-
Legionella Persistence in Manufactured Water Systems: Pasteurization Potentially Selecting for Thermal Tolerance.Front Microbiol. 2017 Jul 19;8:1330. doi: 10.3389/fmicb.2017.01330. eCollection 2017. Front Microbiol. 2017. PMID: 28769899 Free PMC article. Review.
-
Legionella pneumophila: population genetics, phylogeny and genomics.Infect Genet Evol. 2009 Sep;9(5):727-39. doi: 10.1016/j.meegid.2009.05.004. Epub 2009 May 18. Infect Genet Evol. 2009. PMID: 19450709 Review.
References
-
- Allegra S, Grattard F, Girardot F, Riffard S, Pozzetto B, Berthelot P. Longitudinal evaluation of the efficacy of heat treatment procedures against Legionella spp. in hospital water systems by using a flow cytometric assay. Applied and Environmental Microbiology. 2011;77(4):1268–1275. doi: 10.1128/AEM.02225-10. - DOI - PMC - PubMed
-
- Almeida D, Cristovam E, Caldeira D, Ferreira JJ, Marques T. Are there effective interventions to prevent hospital-acquired Legionnaires’ disease or to reduce environmental reservoirs of Legionella in hospitals? A systematic review. American Journal of Infection Control. 2016;44(11):e183–e188. doi: 10.1016/j.ajic.2016.06.018. - DOI - PubMed
-
- Alves LPS, Almeida AT, Cruz LM, Pedrosa FO, de Souza EM, Chubatsu LS, Müller-Santos M, Valdameri G. A simple and efficient method for poly-3-hydroxybutyrate quantification in diazotrophic bacteria within 5 minutes using flow cytometry. Brazilian Journal of Medical and Biological Research. 2017;50(1):e5492. doi: 10.1590/1414-431x20165492. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
