

Repression of developmental transcription factor networks triggers aging-associated gene expression in human glial progenitor cells
- PMID: 38719882
- PMCID: PMC11079006
- DOI: 10.1038/s41467-024-48118-2
Repression of developmental transcription factor networks triggers aging-associated gene expression in human glial progenitor cells
Abstract
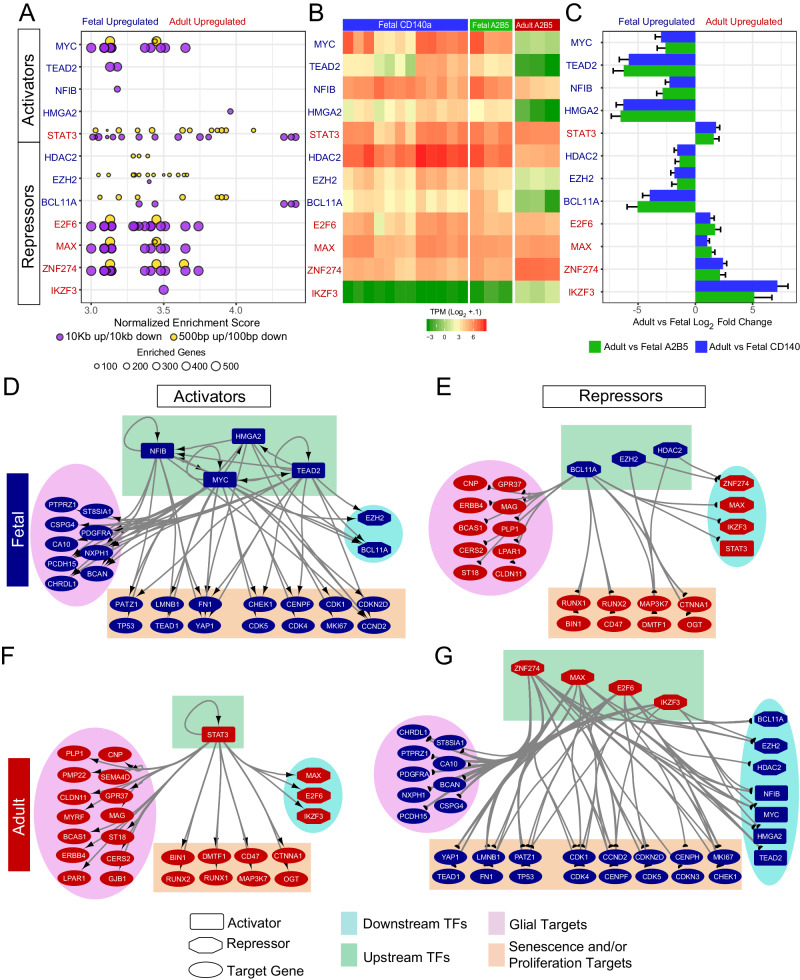
Human glial progenitor cells (hGPCs) exhibit diminished expansion competence with age, as well as after recurrent demyelination. Using RNA-sequencing to compare the gene expression of fetal and adult hGPCs, we identify age-related changes in transcription consistent with the repression of genes enabling mitotic expansion, concurrent with the onset of aging-associated transcriptional programs. Adult hGPCs develop a repressive transcription factor network centered on MYC, and regulated by ZNF274, MAX, IKZF3, and E2F6. Individual over-expression of these factors in iPSC-derived hGPCs lead to a loss of proliferative gene expression and an induction of mitotic senescence, replicating the transcriptional changes incurred during glial aging. miRNA profiling identifies the appearance of an adult-selective miRNA signature, imposing further constraints on the expansion competence of aged GPCs. hGPC aging is thus associated with acquisition of a MYC-repressive environment, suggesting that suppression of these repressors of glial expansion may permit the rejuvenation of aged hGPCs.
© 2024. The Author(s).
Conflict of interest statement
Dr. Goldman is also a part-time employee and stockholder of Sana Biotechnology (Seattle, WA), and his lab receives sponsored research support for projects unrelated to the present work from Sana. Dr. Goldman is also a stockholder and SAB member of CNS2, Inc.; his lab also receives research support from CNS2. N.P.T.H is now a Sana employee, and D.C.M. is a consultant to Sana, in both cases for unrelated work. None of the other authors have any known potential conflicts of interest with regards to this work.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
