

Physiological aging and inflammation-induced cellular senescence may contribute to oligodendroglial dysfunction in MS
- PMID: 38722375
- PMCID: PMC11082024
- DOI: 10.1007/s00401-024-02733-x
Physiological aging and inflammation-induced cellular senescence may contribute to oligodendroglial dysfunction in MS
Abstract
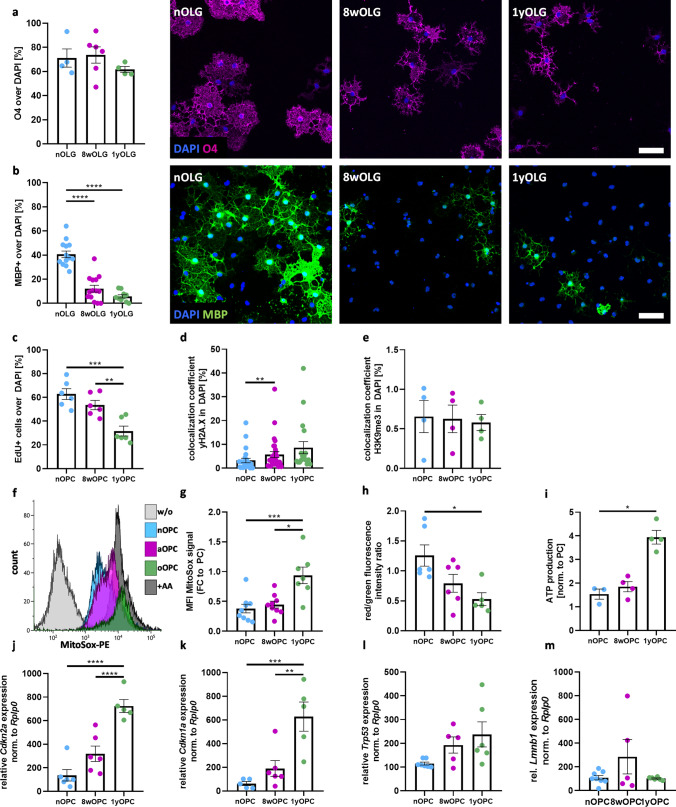
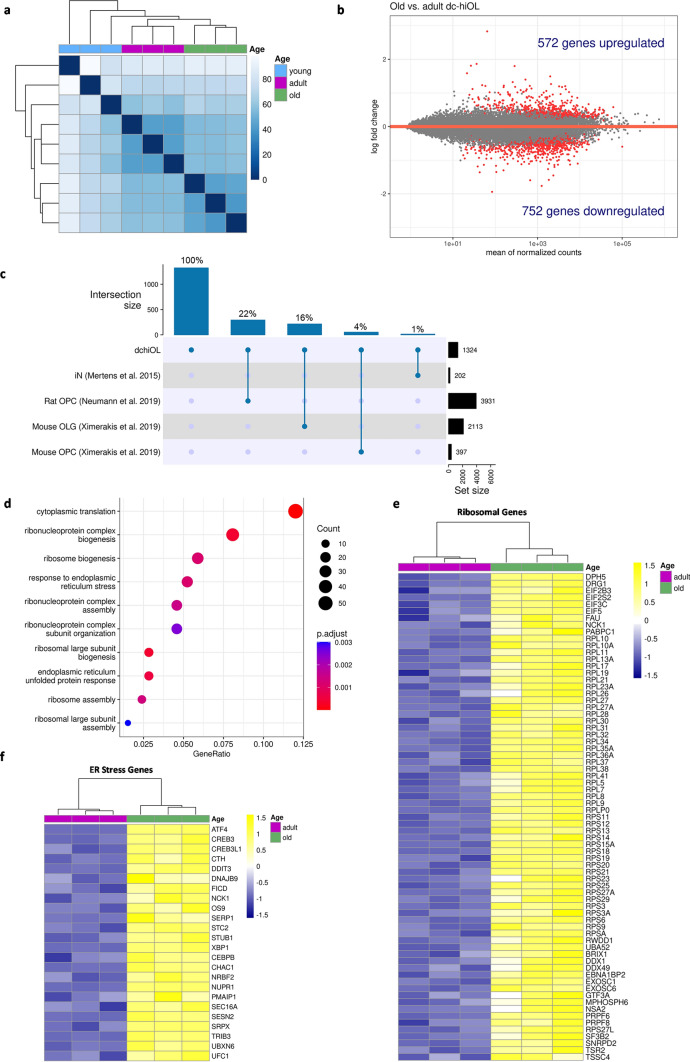
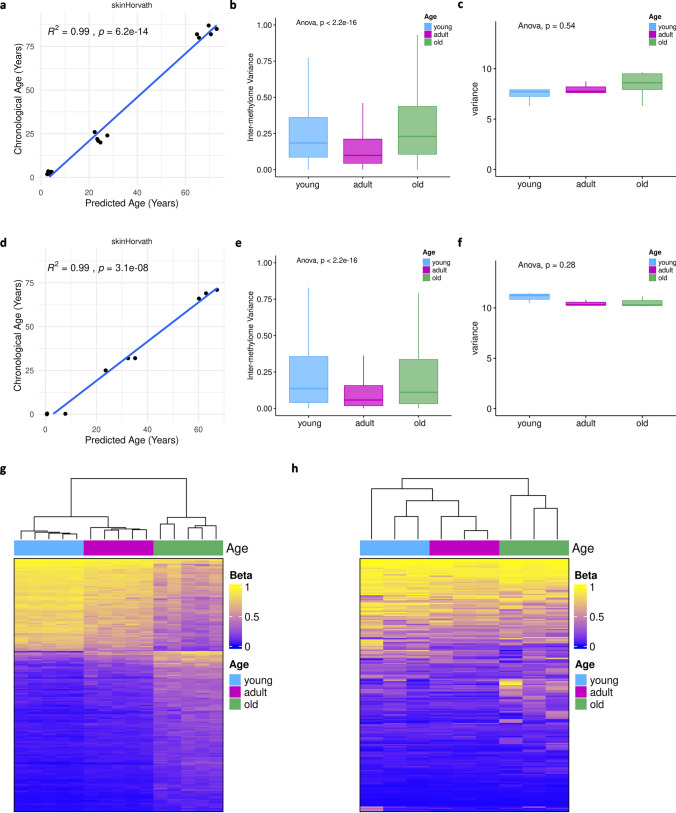
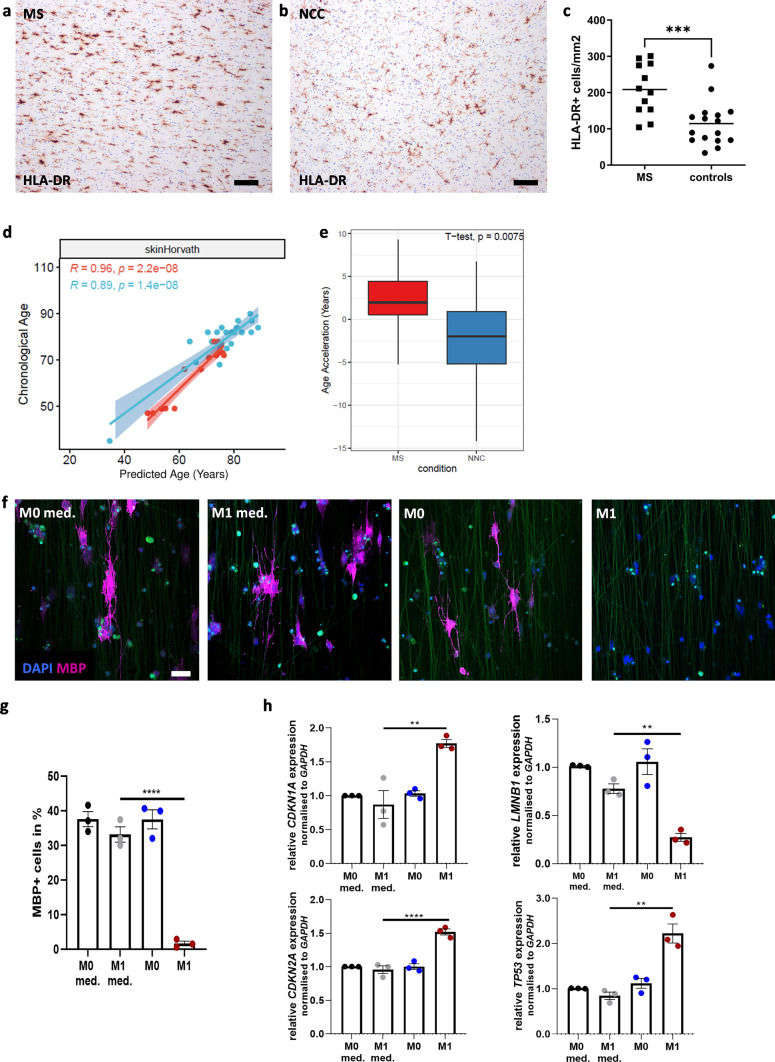
Aging affects all cell types in the CNS and plays an important role in CNS diseases. However, the underlying molecular mechanisms driving these age-associated changes and their contribution to diseases are only poorly understood. The white matter in the aging brain as well as in diseases, such as Multiple sclerosis is characterized by subtle abnormalities in myelin sheaths and paranodes, suggesting that oligodendrocytes, the myelin-maintaining cells of the CNS, lose the capacity to preserve a proper myelin structure and potentially function in age and certain diseases. Here, we made use of directly converted oligodendrocytes (dchiOL) from young, adult and old human donors to study age-associated changes. dchiOL from all three age groups differentiated in an comparable manner into O4 + immature oligodendrocytes, but the proportion of MBP + mature dchiOL decreased with increasing donor age. This was associated with an increased ROS production and upregulation of cellular senescence markers such as CDKN1A, CDKN2A in old dchiOL. Comparison of the transcriptomic profiles of dchiOL from adult and old donors revealed 1324 differentially regulated genes with limited overlap with transcriptomic profiles of the donors' fibroblasts or published data sets from directly converted human neurons or primary rodent oligodendroglial lineage cells. Methylome analyses of dchiOL and human white matter tissue samples demonstrate that chronological and epigenetic age correlate in CNS white matter as well as in dchiOL and resulted in the identification of an age-specific epigenetic signature. Furthermore, we observed an accelerated epigenetic aging of the myelinated, normal appearing white matter of multiple sclerosis (MS) patients compared to healthy individuals. Impaired differentiation and upregulation of cellular senescence markers could be induced in young dchiOL in vitro using supernatants from pro-inflammatory microglia. In summary, our data suggest that physiological aging as well as inflammation-induced cellular senescence contribute to oligodendroglial pathology in inflammatory demyelinating diseases such as MS.
Keywords: Aging; Direct conversion; Human oligodendrocytes; Multiple sclerosis.
© 2024. The Author(s).
Conflict of interest statement
TK received research funding from the German Research Foundation, Interdisciplinary Center for Clinical Research (IZKF) Münster, National MS Society, German MS Society and Novartis. She received compensation for serving on scientific advisory boards (Novartis, Sanofi, Merck) and speaker honoraria from Novartis, Biogen and Roche.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
