

Behavioral control through the direct, focal silencing of neuronal activity
- PMID: 38729162
- PMCID: PMC11260259
- DOI: 10.1016/j.chembiol.2024.04.003
Behavioral control through the direct, focal silencing of neuronal activity
Abstract
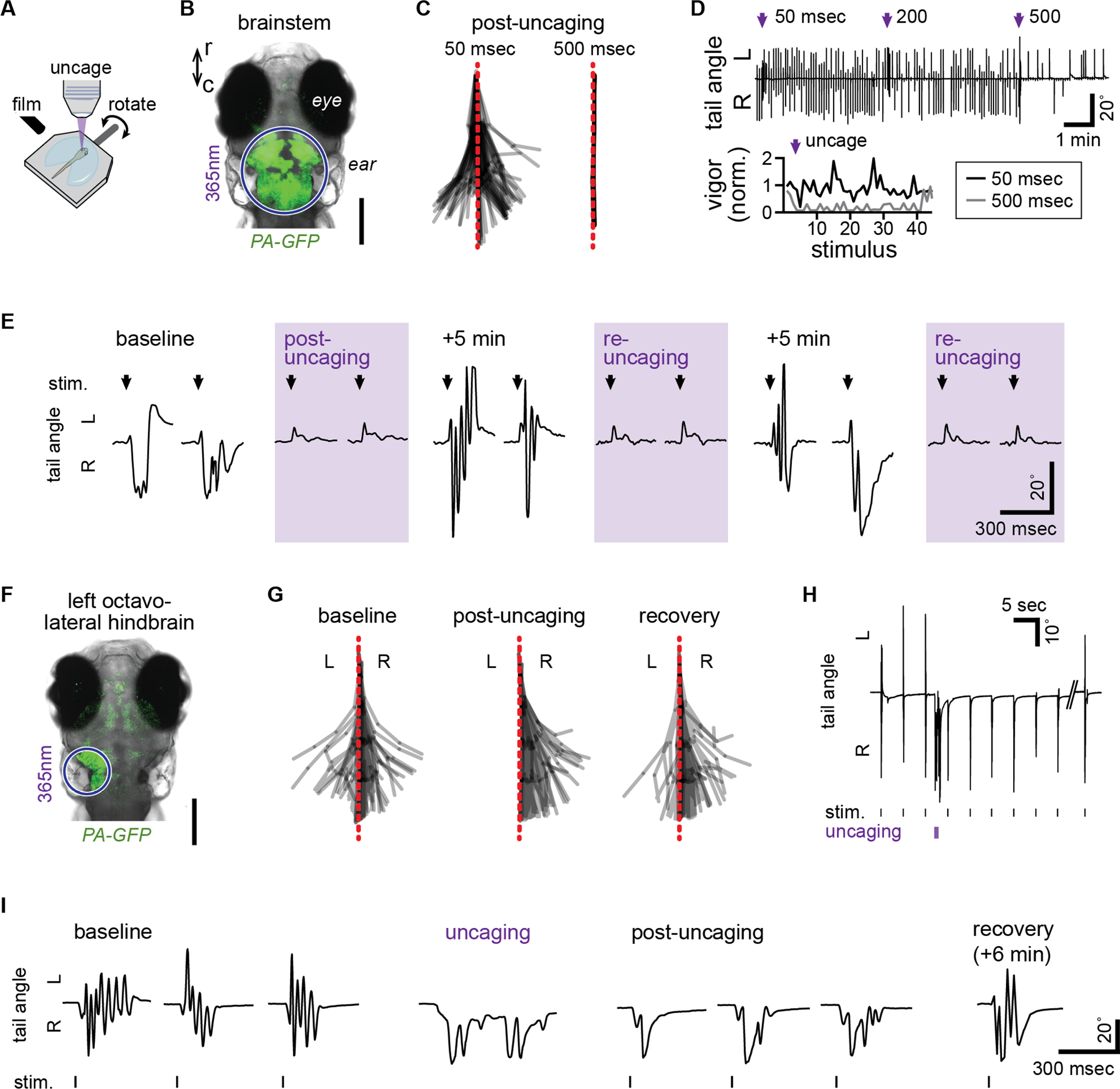
The ability to optically stimulate and inhibit neurons has revolutionized neuroscience research. Here, we present a direct, potent, user-friendly chemical approach for optically silencing neurons. We have rendered saxitoxin (STX), a naturally occurring paralytic agent, transiently inert through chemical protection with a previously undisclosed nitrobenzyl-derived photocleavable group. Exposing the caged toxin, STX-bpc, to a brief (5 ms) pulse of light effects rapid release of a potent STX derivative and transient, spatially precise blockade of voltage-gated sodium channels (NaVs). We demonstrate the efficacy of STX-bpc for parametrically manipulating action potentials in mammalian neurons and brain slice. Additionally, we show the effectiveness of this reagent for silencing neural activity by dissecting sensory-evoked swimming in larval zebrafish. Photo-uncaging of STX-bpc is a straightforward method for non-invasive, reversible, spatiotemporally precise neural silencing without the need for genetic access, thus removing barriers for comparative research.
Keywords: actional potential; electrophysiology; photocage; saxitoxin; sodium channel; zebrafish.
Copyright © 2024 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests J.D. is a cofounder and holds equity shares in SiteOne Therapeutics, Inc., a start-up company interested in developing subtype-selective Na(V) modulators.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
