

A time-resolved multi-omics atlas of transcriptional regulation in response to high-altitude hypoxia across whole-body tissues
- PMID: 38730227
- PMCID: PMC11087590
- DOI: 10.1038/s41467-024-48261-w
A time-resolved multi-omics atlas of transcriptional regulation in response to high-altitude hypoxia across whole-body tissues
Abstract
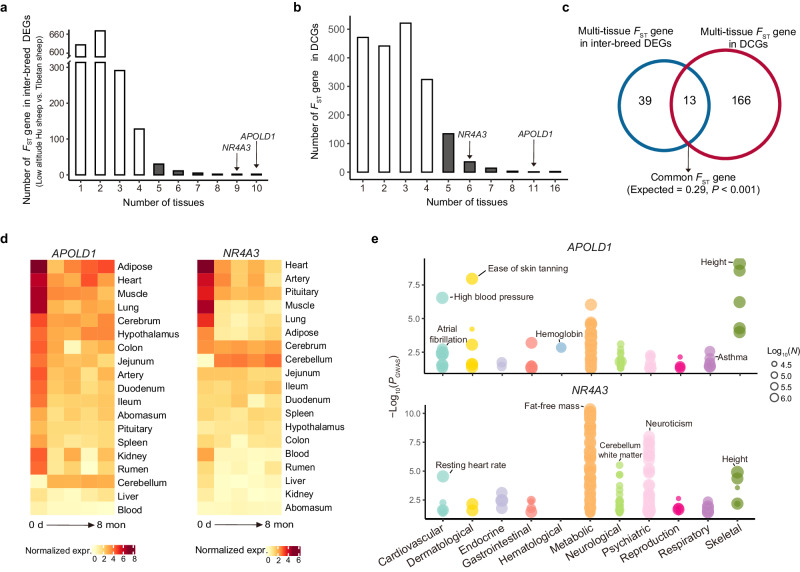
High-altitude hypoxia acclimatization requires whole-body physiological regulation in highland immigrants, but the underlying genetic mechanism has not been clarified. Here we use sheep as an animal model for low-to-high altitude translocation. We generate multi-omics data including whole-genome sequences, time-resolved bulk RNA-Seq, ATAC-Seq and single-cell RNA-Seq from multiple tissues as well as phenotypic data from 20 bio-indicators. We characterize transcriptional changes of all genes in each tissue, and examine multi-tissue temporal dynamics and transcriptional interactions among genes. Particularly, we identify critical functional genes regulating the short response to hypoxia in each tissue (e.g., PARG in the cerebellum and HMOX1 in the colon). We further identify TAD-constrained cis-regulatory elements, which suppress the transcriptional activity of most genes under hypoxia. Phenotypic and transcriptional evidence indicate that antenatal hypoxia could improve hypoxia tolerance in offspring. Furthermore, we provide time-series expression data of candidate genes associated with human mountain sickness (e.g., BMPR2) and high-altitude adaptation (e.g., HIF1A). Our study provides valuable resources and insights for future hypoxia-related studies in mammals.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
