

Design of antiviral AGO2-dependent short hairpin RNAs
- PMID: 38734183
- PMCID: PMC11401469
- DOI: 10.1016/j.virs.2024.05.001
Design of antiviral AGO2-dependent short hairpin RNAs
Abstract
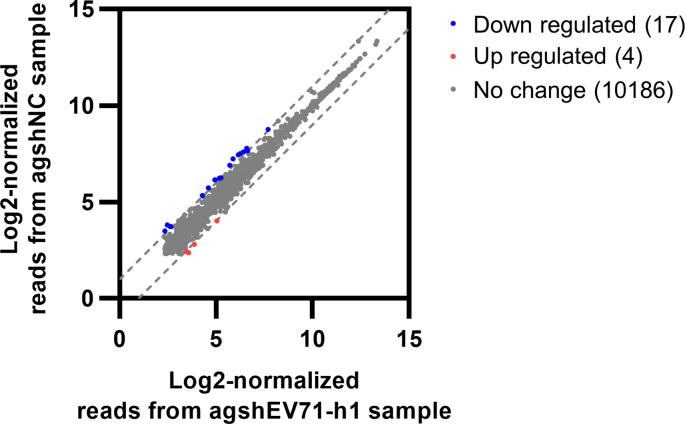
The increasing emergence and re-emergence of RNA virus outbreaks underlines the urgent need to develop effective antivirals. RNA interference (RNAi) is a sequence-specific gene silencing mechanism that is triggered by small interfering RNAs (siRNAs) or short hairpin RNAs (shRNAs), which exhibits significant promise for antiviral therapy. AGO2-dependent shRNA (agshRNA) generates a single-stranded guide RNA and presents significant advantages over traditional siRNA and shRNA. In this study, we applied a logistic regression algorithm to a previously published chemically siRNA efficacy dataset and built a machine learning-based model with high predictive power. Using this model, we designed siRNA sequences targeting diverse RNA viruses, including human enterovirus A71 (EV71), Zika virus (ZIKV), dengue virus 2 (DENV2), mouse hepatitis virus (MHV) and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), and transformed them into agshRNAs. We validated the performance of our agshRNA design by evaluating antiviral efficacies of agshRNAs in cells infected with different viruses. Using the agshRNA targeting EV71 as an example, we showed that the anti-EV71 effect of agshRNA was more potent compared with the corresponding siRNA and shRNA. Moreover, the antiviral effect of agshRNA is dependent on AGO2-processed guide RNA, which can load into the RNA-induced silencing complex (RISC). We also confirmed the antiviral effect of agshRNA in vivo. Together, this work develops a novel antiviral strategy that combines machine learning-based algorithm with agshRNA design to custom design antiviral agshRNAs with high efficiency.
Keywords: AgshRNA; Antiviral strategy; Machine learning model.
Copyright © 2024 The Authors. Publishing services by Elsevier B.V. All rights reserved.
Conflict of interest statement
Conflict of interest Professor Xi Zhou is an editorial board member for Virologica Sinica and were not involved in the editorial review or the decision to publish this article. The authors declare that they have no conflict of interest.
Figures





References
-
- Anobile D.P., Poirier E.Z. RNA interference, an emerging component of antiviral immunity in mammals. Biochem. Soc. Trans. 2023;51:137–146. - PubMed
-
- Birmingham A., Anderson E.M., Reynolds A., Ilsley-Tyree D., Leake D., Fedorov Y., Baskerville S., Maksimova E., Robinson K., Karpilow J., Marshall W.S., Khvorova A. 3’ utr seed matches, but not overall identity, are associated with RNAi off-targets. Nat. Methods. 2006;3:199–204. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
