

A commentary on studies of brain iron accumulation during ageing
- PMID: 38735007
- PMCID: PMC11186910
- DOI: 10.1007/s00775-024-02060-2
A commentary on studies of brain iron accumulation during ageing
Abstract
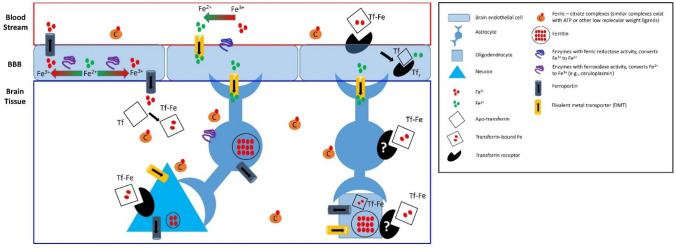
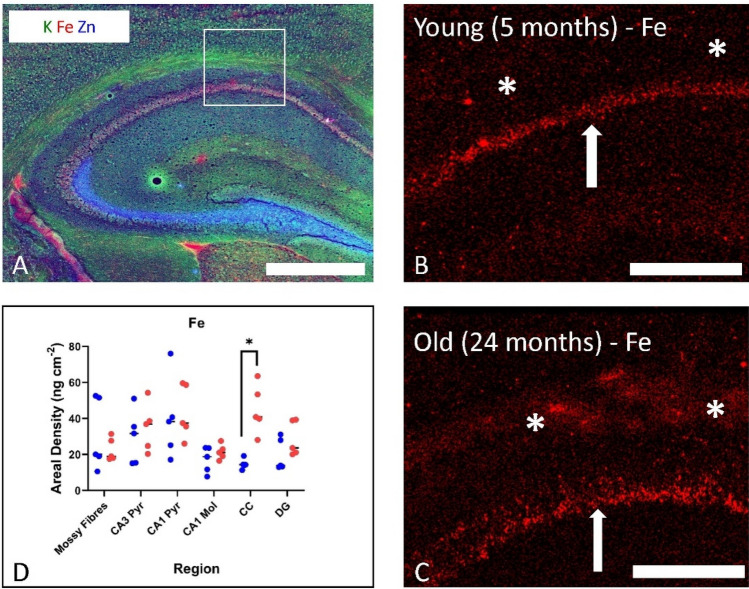
Brain iron content is widely reported to increase during "ageing", across multiple species from nematodes, rodents (mice and rats) and humans. Given the redox-active properties of iron, there has been a large research focus on iron-mediated oxidative stress as a contributor to tissue damage during natural ageing, and also as a risk factor for neurodegenerative disease. Surprisingly, however, the majority of published studies have not investigated brain iron homeostasis during the biological time period of senescence, and thus knowledge of how brain homeostasis changes during this critical stage of life largely remains unknown. This commentary examines the literature published on the topic of brain iron homeostasis during ageing, providing a critique on limitations of currently used experimental designs. The commentary also aims to highlight that although much research attention has been given to iron accumulation or iron overload as a pathological feature of ageing, there is evidence to support functional iron deficiency may exist, and this should not be overlooked in studies of ageing or neurodegenerative disease.
Keywords: Brain rust; Metallomics; Senescence; X-ray fluorescence; XFM.
© 2024. The Author(s).
Conflict of interest statement
MJH has no conflicts of interest to declare.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
