

Protective role of RIPK1 scaffolding against HDV-induced hepatocyte cell death and the significance of cytokines in mice
- PMID: 38739648
- PMCID: PMC11115361
- DOI: 10.1371/journal.ppat.1011749
Protective role of RIPK1 scaffolding against HDV-induced hepatocyte cell death and the significance of cytokines in mice
Abstract
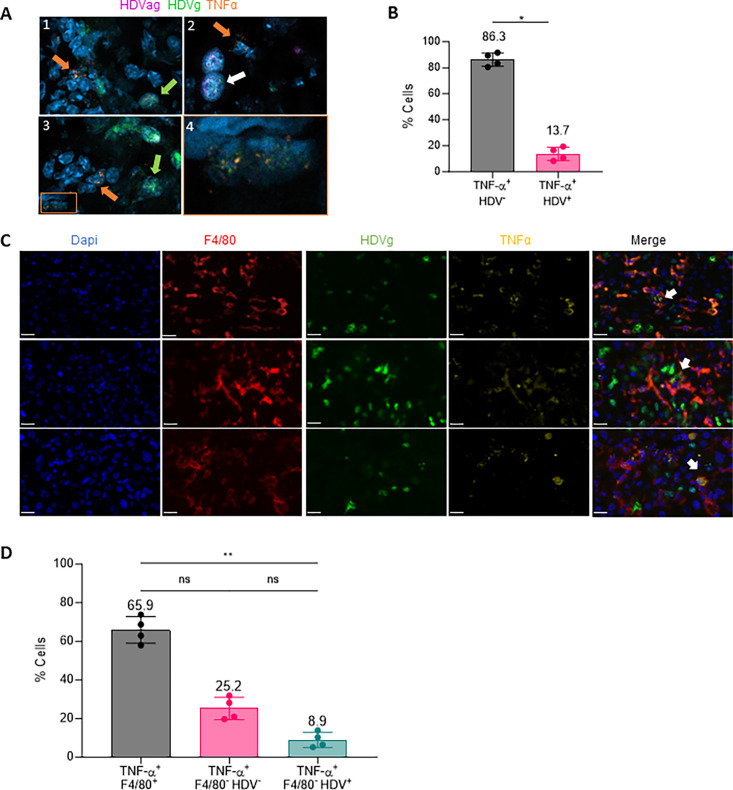
Hepatitis delta virus (HDV) infection represents the most severe form of human viral hepatitis; however, the mechanisms underlying its pathology remain incompletely understood. We recently developed an HDV mouse model by injecting adeno-associated viral vectors (AAV) containing replication-competent HBV and HDV genomes. This model replicates many features of human infection, including liver injury. Notably, the extent of liver damage can be diminished with anti-TNF-α treatment. Here, we found that TNF-α is mainly produced by macrophages. Downstream of the TNF-α receptor (TNFR), the receptor-interacting serine/threonine-protein kinase 1 (RIPK1) serves as a cell fate regulator, playing roles in both cell survival and death pathways. In this study, we explored the function of RIPK1 and other host factors in HDV-induced cell death. We determined that the scaffolding function of RIPK1, and not its kinase activity, offers partial protection against HDV-induced apoptosis. A reduction in RIPK1 expression in hepatocytes through CRISPR-Cas9-mediated gene editing significantly intensifies HDV-induced damage. Contrary to our expectations, the protective effect of RIPK1 was not linked to TNF-α or macrophage activation, as their absence did not alter the extent of damage. Intriguingly, in the absence of RIPK1, macrophages confer a protective role. However, in animals unresponsive to type-I IFNs, RIPK1 downregulation did not exacerbate the damage, suggesting RIPK1's role in shielding hepatocytes from type-I IFN-induced cell death. Interestingly, while the damage extent is similar between IFNα/βR KO and wild type mice in terms of transaminase elevation, their cell death mechanisms differ. In conclusion, our findings reveal that HDV-induced type-I IFN production is central to inducing hepatocyte death, and RIPK1's scaffolding function offers protective benefits. Thus, type-I IFN together with TNF-α, contribute to HDV-induced liver damage. These insights may guide the development of novel therapeutic strategies to mitigate HDV-induced liver damage and halt disease progression.
Copyright: © 2024 Camps et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
